Avimimus
| Avimimus Temporal range: Late Cretaceous, 70 Ma | |
|---|---|
 | |
| Reconstructed skeleton | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Reptilia |
| Clade: | Dinosauria |
| Order: | Saurischia |
| Suborder: | Theropoda |
| Superfamily: | †Caenagnathoidea |
| Family: | †Avimimidae Kurzanov, 1981 |
| Genus: | †Avimimus Kurzanov, 1981 |
| Species: | †A. portentosus |
| Binomial name | |
| Avimimus portentosus Kurzanov, 1981 | |
Avimimus (/ˌeɪvᵻˈmaɪməs/ AY-vi-MY-məs), meaning "bird mimic" (Latin avis = bird + mimus = mimic), was a genus of oviraptorosaurian therapod dinosaur, named for its bird-like characteristics, that lived in the late Cretaceous in what is now Mongolia, around 70 million years ago.
Description

Avimimus was a small, bird-like dinosaur with a length of 1.5 m (5 ft).[1] The skull was small compared to the body, though the brain[1] and eyes were large. The size of the bones which surrounded the brain and were dedicated to protecting it are large.[1] This is also consistent with the hypothesis that Avimimus had a proportionally large brain.[1]
The jaws of Avimimus were thought to form a parrot-like beak lacking teeth, and a thorough review of the holotype specimen's anatomy confirmed that no teeth were preserved, although a series of tooth-like projections along the tip of the premaxilla were.[2] However, subsequently discovered specimens have been reported to preserve small premaxillary teeth.[3] The small teeth or possible lack thereof in Avimimus suggests that it may have been an herbivore or omnivore. Kurzanov himself, however, believed that Avimimus was an insectivore.[1]
The foramen magnum, the hole allowing the spinal cord to connect with the brain, was proportionally large in Avimimus.[1] The occipital condyle, however, was small, further suggestive of the skull's relative lightness.[1] The neck itself was long and slender, and is composed of vertebrae that are much more elongate than in other oviraptorosaurs. Unlike oviraptorids and caenagnathids, the back vertebrae lack openings for air sacs, suggesting that Avimimus is more primitive than these animals.
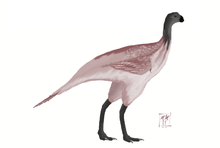
The forelimbs were relatively short. The bones of the hand were fused together, as in modern birds, and a ridge on the ulna (lower arm bone) was interpreted as an attachment point for feathers by Kurzanov.[1][4] Kurzanov, in 1987, also reported the presence of quill knobs,[1][5] and while Chiappe confirmed the presence of bumps on the ulna, their function remained unclear.[2] Kurzanov was so convinced they were attachment points for feathers that he concluded that Avimimus may have been capable of weak flight.[1] The presence of feathers is now widely accepted, but most paleontologists do not believe Avimimus could fly.[1]
The ilium was almost horizontally oriented, resulting in exceptionally broad hips. Little is known of the tail but the hip suggests that the tail was long. The legs were extremely long and slender, suggesting that Avimimus was a highly specialized runner. The proportions of the leg bones add further weight to the idea of Avimimus was quick on its feet.[1] The animal's shins were long in comparison with its thighs,[1] a trait common among cursorial animals. It also had three-toed feet with narrow pointed claws.
Discovery and species
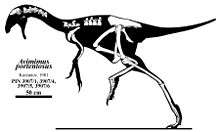
The remains of Avimimus were recovered by Russian paleontologists[1] and officially described by Dr. Sergei Kurzanov in 1981. The Avimimus fossils were initially described as having come from the Djadokta Formation by Kurzanov; however, in a 2006 description of a new specimen, Watabe and colleagues noted that Kurzanov was probably mistaken about the provenance, and it is more likely that Avimimus hailed from the more recent Nemegt Formation.[6] The type species is A. portentosus.[4] Because no tail was found with the original find, Kurzanov mistakenly concluded that Avimimus lacked a tail in life.[1] However, subsequent Avimimus finds containing caudal vertebrae have confirmed the presence of a tail.[1] A second nearly complete specimen of Avimimus was discovered in 1996 and described in 2000 by Watabe and colleagues. Additionally, these authors identified a number of small theropod footprints in the same area as belonging to Avimimus.[3]

A variety of isolated bones that have been attributed to Avimimus but differ from A. portentosus may represent one or more distinct species, referred to as Avimimus sp..[7]
In 2008, a team of Canadian, American, and Mongolian paleontologists headed by Phil Currie reported the discovery of an extensive bonebed of Avimimus sp. fossils. The bonebed is in the Nemegt Formation, 10.5 meters above the Barun Goyot Formation, in the Gobi Desert. The team reported finding abundant bones of at least ten individuals of Avimimus, but the deposit may hold more. All individuals were either adult or subadult, and the adults showed little variation in size, suggesting determinate growth. The team also suggests that the individuals were found together because they were gregarious in life, providing possible indications that Avimimus formed age-segregated groups for either lekking or flocking purposes. The adults showed a greater degree of skeletal fusion in the tarsometatarsus and tibiotarsus, and also more prominent muscle scars. The preservation of the bonebeds suggest that they were buried rapidly, uncovered by rapid flow of water, and then buried again a short distance away.[8][9]
Classification
Avimimus was originally suggested to be a very close relative of birds, given its unique suite of bird-like features not known in other dinosaurs at the time. In fact, Kurzanov argued that Avimimus, rather than the famous early bird Archaeopteryx, was close to the direct ancestor of modern birds, and that Archaeopteryx was not as closely related to birds as had previously been suggested.[5] However, this view has not been supported by later phylogenetic analyses of dinosaur and bird relationships. Most modern scientists find that Avimimus in fact belongs to a diverse group of bird-like dinosaurs more primitive than Archaeopteryx, the oviraptorosaurs.[10][11]
Kurzanov placed Avimimus in its own family, Avimimidae, in 1981. In 1991, Sankar Chatterjee erected the Order Avimimiformes to include Avimimus. Neither of these group names is used frequently by paleontologists as they include only a single species. More recent studies have shown that Avimimus is best grouped within the Oviraptoridae, within the subgroup Elmisaurinae.[12]
The following cladogram follows an analysis by Phil Senter, 2007.[13]
| Caenagnathoidea |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
Paleoecology
The Barun Goyot Formation of Mongolia, is estimated to date back to the Campanian stage, between 84 and 70 million years ago[14] of the Late Cretaceous period.[15] During the Late Cretaceous period, the land that is now the Barun Goyot Formation had an arid environment with fields of sand dunes and only intermittent streams. It is slightly younger than the nearby Djadochta Formation, and seems to have been slightly wetter.[15] This formation is noteworthy for the exquisite preservation of small and fragile dinosaur skeletons, a rare occurrence considering that these fossils are typically broken up and dispersed when found in other rock formations.[16]
The region that is preserved in the Barun Goyot Formation was home to the maniraptoran Hulsanpes perlei, the oviraptorids Conchoraptor gracilis and Ajancingenia yanshini, the alvarezsaurids Ceratonykus oculatus, Mononykus and Parvicursor remotus, the pachycephalosaur Tylocephale gilmorei, the ankylosaurs Saichania chulsanensis and Tarchia gigantea, and the ceratopsians Bagaceratops rozhdestvenskyi, Breviceratops kozlowskii, Lamaceratops tereschenkoi and Platyceratops tatarinovi. The largest dinosaur among them was the titanosaur Quaesitosaurus orientalis. It was observed that many of the same genera were present at the Barun Goyot and Djadochta Formations, though there was variation at the species level.[17] Vertebrates present in the Barun Goyot Formation included the primitive birds Gobipteryx minuta and Hollanda luceria and the lizards Estesia mongoliensis, Ovoo gurvel, Proplatynotia longirostrata and Gobiderma pulchrum. The early mammals that were present in this region during the time of Avimimus were the placental mammals Asioryctes nemegetensis and Barunlestes butleri, the amphibian Gobiates khermeentsavi, the multituberculate mammals Catopsbaatar catopsaloides, Chulsanbaatar vulgaris and Nemegtbaatar gobiensis, and the marsupial mammal Asiatherium reshetovi and Deltatheridium pretrituberculare. Numerous dinosaurs eggshells found in this region support the presence of sauropods and maniraptors.
See also
References
- 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 "Avimimus." In: Dodson, Peter & Britt, Brooks & Carpenter, Kenneth & Forster, Catherine A. & Gillette, David D. & Norell, Mark A. & Olshevsky, George & Parrish, J. Michael & Weishampel, David B. The Age of Dinosaurs. Publications International, LTD. p. 130. ISBN 0-7853-0443-6.
- 1 2 Chiappe, L.M. and Witmer, L.M. (2002). Mesozoic Birds: Above the Heads of Dinosaurs. Berkeley: University of California Press, 536 pp. ISBN 0-520-20094-2
- 1 2 Watabe, Weishampel, Barsbold, Tsogtbaatar and Suzuke, 2000. "New nearly complete skeleton of the bird-like theropod, Avimimus, from the Upper Cretaceous of the Gobi Desert, Mongolia." Journal of Vertebrate Paleontology, 20(3): 77A.
- 1 2 Kurzanov, S.M. (1981). "An unusual theropod from the Upper Cretaceous of Mongolia Iskopayemyye pozvonochnyye Mongolii (Fossil Vertebrates of Mongolia)." Trudy Sovmestnay Sovetsko-Mongolskay Paleontologiyeskay Ekspeditsiy (Joint Soviet-Mongolian Paleontological Expedition), 15: 39-49. Nauka Moscow, 1981
- 1 2 Kurzanov, S.M. (1987). "Avimimidae and the problem of the origin of birds." Transactions of the Joint Soviet-Mongolian Paleontological Expedition, 31: 5-92. [in Russian]
- ↑ Watabe, Suzuki and Tsogtbaatar, 2006. Geological and geographical distribution of bird-like theropod, Avimimus in Mongolia. Journal of Vertebrate Paleontology. 26(3), 136A-137A.
- ↑ Ryan, Currie, P. and Russell, D. (2001). "New material of Avimimus portentosus (Theropoda) from the Iren Debasu Formation (Upper Cretaceous) of the Erenhot Region of Inner Mongolia." Journal of Vertebrate Paleontology, 21(3): 95A.
- ↑ Currie, P.; Longrich, N.; Ryan, M.; Eberth, D.; Demchig, B. (2008). "A bonebed of Avimimus sp. (Dinosauria: Theropoda) from the Late Cretaceous Nemegt Formation, Gobi Desert: Insights into social behavior and development in a maniraptoran theropod.". Journal of Vertebrate Paleontology. 28 (3): 67A.
- ↑ Funston, G.F.; Currie, P.J.; Eberth, D.A.; Ryan, M.J.; Chinzorig, T.; Badamgarav, D.; Longrich, N.R. (2016). "The first oviraptorosaur (Dinosauria: Theropoda) bonebed: evidence of gregarious behaviour in a maniraptoran theropod". Scientific Reports. 6: 35782. doi:10.1038/srep35782.
- ↑ Dyke and Thorley (1998). "Reduced cladistic consensus methods and the avian affinities of Protoavis and Avimimus." Archaeopteryx, 16: 123-129.
- ↑ Turner, Alan H.; Pol, Diego; Clarke, Julia A.; Erickson, Gregory M.; Norell, Mark (2007). "A basal dromaeosaurid and size evolution preceding avian flight" (pdf). Science. 317 (5843): 1378–1381. doi:10.1126/science.1144066. PMID 17823350.
- ↑ Holtz, Thomas R. Jr. (2010) Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages, Winter 2010 Appendix.
- ↑ Senter, P. (2007). "A new look at the phylogeny of Coelurosauria (Dinosauria: Theropoda)." Journal of Systematic Palaeontology, (doi:10.1017/S1477201907002143).
- ↑ Gradstein, Felix M.; Ogg, James G.; Smith, Alan G. (2005). A Geologic Time Scale 2004. Cambridge: Cambridge University Press. ISBN 978-0-521-78142-8.
- 1 2 Jerzykiewicz, Tomasz; Russell, Dale A. (1991). "Late Mesozoic stratigraphy and vertebrates of the Gobi Basin". Cretaceous Research. 12 (4): 345–377. doi:10.1016/0195-6671(91)90015-5.
- ↑ Novacek, M. J., 1996. "Dinosaurs of the Flaming. Cliffs" Anchor/Doubleday. 367 p.
- ↑ Nicholas R. Longrich; Philip J. Currie; Dong Zhi-Ming (2010). "A new oviraptorid (Dinosauria: Theropoda) from the Upper Cretaceous of Bayan Mandahu, Inner Mongolia". Palaeontology. 53 (5): 945–960. doi:10.1111/j.1475-4983.2010.00968.x.