Cycle (gene)
| Cycle | |
|---|---|
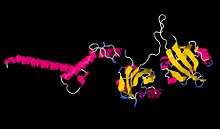 The approximate 3D structure of CYC protein generated with Phyre2. The protein sequence information was obtained from the UniProt database. | |
| Identifiers | |
| Organism | |
| Symbol | cyc |
| Alt. symbols | bmal1 |
| Entrez | 40162 |
| RefSeq (mRNA) | NM_079444.3 |
| RefSeq (Prot) | NP_524168.2 |
| UniProt | O61734 |
| Other data | |
| Chromosome | 3L: 19.81 - 19.81 Mb |
| aryl hydrocarbon receptor nuclear translocator-like | |
|---|---|
| Identifiers | |
| Symbol | ARNTL |
| Alt. symbols | bmal1 |
| Entrez | 406 |
| HUGO | 701 |
| OMIM | 602550 |
| RefSeq | NP_001025443 |
| UniProt | O00327 |
| Other data | |
| Locus | Chr. 11 p15 |
Cycle (cyc) is a gene in Drosophila melanogaster that encodes the CYCLE protein. The Cycle gene is expressed in a variety of cell types in a circadian manner. It is involved in controlling both the sleep-wake cycle and circadian regulation of gene expression by promoting transcription in a negative feedback mechanism. The cyc gene is located on the left arm of chromosome 3 and codes for a transcription factor containing a basic helix-loop-helix (bHLH) domain and a PAS domain.[1] The cyc gene is 2.17 kb in size, made up of 5 coding exons totaling 1,625 base pairs coding for 413 aminos acid residues, and there are currently 19 known alleles.[2][3] Orthologs performing the same function in other species include ARNTL and BMAL1.
Function
Cyc is primarily known for its role in the genetic transcription-translation feedback loop that generates circadian rhythms in Drosophila. In the cell nucleus, the CYCLE protein (CYC) forms a heterodimer with a second bHLH-PAS protein, CLOCK (CLK). This CYC-CLK protein complex binds to E-box elements in promoter regions of the genes period and timeless, functioning as a transcription factor in the translation of the proteins PER and TIM.[4] After PER and TIM proteins accumulate in the cytoplasm and bind together, the PER-TIM complex translocates to the nucleus and interferes with the CYC-CLK protein complex function to inhibit its activation of transcription. Thus, CLK and CYC act as positive factors and PER and TIM as negative factors. CYC also plays a role in the post-translational regulation of CLK in the cytoplasm.[5] These four proteins of the feedback loop are later degraded by a casein kinase-mediated phosphorylation cycle, allowing fluctuations in gene expression according to environmental cues. The Howard Hughes Medical Institute has an educational video detailing the transcription-translation feedback loop.
Cyc is involved with the genetic basis of other behaviors that relate to circadian processes, such as Drosophila’s responses to starvation and sleep deprivation, and regulation of rest and life span. Starvation in Drosophila potently suppresses sleep, suggesting that the homeostatically regulated behaviors of feeding and sleep are integrated in flies. Clk and cyc act during starvation to modulate the conflict of whether flies sleep or search for food, thus playing a critical role for proper sleep suppression during starvation.[6]
The role of cyc in response to sleep deprivation involves expression of heat-shock genes, but is not fully understood.[7] Its role in regulation of rest and longevity in Drosophila is also not well understood, but evidence indicates that this role displays gender dimorphism.
Cyc is expressed constitutively (continuously) in Drosophila cells[4] and is present in naive Drosophila tissue culture cells, unlike clk, per, or tim.[8] Regulation thus occurs primarily through the negative feedback by the PER-TIM protein complex in the transcription-translation feedback loop described above.
Discovery
The identification, characterization, and cloning of cycle was reported in May 1998 in Cell by Jeffrey Hall and Michael Rosbash’s labs at Brandeis University along with first author Joan E. Rutila at the Howard Hughes Medical Institute.[4] Prior to its discovery, the mechanism by which PER and TIM transcription was regulated rhythmically was not fully understood. They published the papers reporting the discovery of CYCLE and CLOCK in the same issue of Cell. They found both genes as a result of chemically mutagenzing flies and screening for altered locomotor activity rhythms.[9] From the screen, cycle was identified as a recessive arrhythmic mutant in one fly line because it shows arrhythmic locomotor activity patterns when a fly has 2 mutant chromosomes number 3.[4] These mutant flies were also found to display arrhythmic eclosion.[4] Because the mutants displayed no circadian rhythms and the heterozygote flies displayed long circadian periods, they determined that cycle has a dominant phenotype. These data also suggest that the Cyclegene is part of the biological clock because of the similarity between the cycle mutant phenotype and that of the clock mutant.[4] This suggests that Cycle is part of the biological clock with its phenotype similar to that of the clock mutant. Assaying PER and TIM transcription levels in the cyc mutant showed reduced mRNA levels of both proteins. Cloning of the cyc gene revealed that it encodes a novel bHLH-PAS protein related to mammalian bmal1, and that it likely binds to Clock to activate transcription of circadian rhythm genes.[4]
Cycle gene expression has been discovered in a variety of cell types and tissues including the adult head, adult eye, larval/adult central nervous system, adult crop, adult midgut, adult hindgut, larval/adult Malpighian tubules, larval/adult fat body, adult salivary gland, adult female reproductive system, adult male accessory gland, and adult carcass.[3]
Recent research on cycle has largely focused on the role of circadian rhythmicity in other processes. In 2012, it was reported that aging reduces transcriptional oscillations of core clock genes in the fly head including cycle.[10] Wild type Drosophila show low activity of the CLOCK/CYCLE protein dimer in the morning, and it was recently found that lowering levels of these proteins can affect neuronal signaling.[11] Research from 2012 on sleep architecture and nutrition found that circadian clock mutants, including cyc01 still maintained a normal diet response without circadian rhythmicity.[12] Future work focusing on understanding the role of circadian rhythms in Drosophila will continue to investigate cycle's role in maintaining rhythmicity.
Species distribution
.jpg)
The cycle gene found in Drosophila melanogaster has many orthologs among eukaryotes including other members of the Drosophila genus, mosquitoes, various non-dipteran insects, non insect arthropods, humans, and other mammals. In other members of the Drosophila genus, functional orthologs of the D. melanogaster cycle gene can either be found in chromosome 3 or in scaffold/matrix attachment regions. In each case, the orthologs retain functional PAS domains, signal transduction function, and transcription factor activity. Other non-arthropods containing the functional ortholog of the Drosophila cycle ARNTL and ARNTL2 include Humans, house mice, domestic chicken and zebrafish. Most vertebrate creatures retain a functionally and structurally similar protein. Unlike dipterans, however, these animals have two different orthologs of the cycle gene most likely caused by a gene duplication event.[13] Much like CYCLE, the ARNTL proteins are basic helix-loop-helix and a PAS domain containing transcription factors responsible for the autoregulatory transcription translation negative feedback loops (above), which are responsible for generating molecular circadian rhythms.[14] For a more complete list of ARNTL homologs visit the ARNTL species distribution article.
Mutations
There are currently 19 known alleles of cyc found in Drosophila melanogaster, and most of these have been mutagenized and engineered by researchers in the laboratory.
Cyc01
Cyc01 also known as cyc0 is a recessive null mutant allele. This means that a Drosophila with two copies of the cyc01 mutant does not produce a functional CYCLE protein. The resulting Drosophila exhibits arrhythmic activity and cannot entrain to any light-dark cycle. Cyc01 mutants showed a disproportionately large sleep rebound and died after 10 hours of sleep deprivation, although they were more resistant than other clock mutants to various stressors. Unlike other clock mutants, cyc01 flies showed a reduced expression of heat-shock genes after sleep loss. However, activating heat-shock genes before sleep deprivation rescued cyc01 flies from its lethal effects.[7]
Cyc02
Cyc02 is a recessive mutant, characterized by a severe reduction in levels of PER protein. In each case, the mutation was the result of a nonsense mutation in the PAS-encoding region found in 1999 following a forward screen of ethyl methanesulfonatemutants. Both the cyc01 and the cyc02 mutants were identified by the same team.[15]
CycΔ
CycΔ mutation is a dominant-negative mutation which blocks the ability of CYCLE-CLOCK complexes from activating E-box dependent transcription of timeless. The mutation is the result of a 15 to 17 base pair deletion from the cyc gene.[16]
cycG4677
A cycG4677 mutant strain is available from Bloomington Drosophila Stock Center at Indiana University. The cycG4677 mutant strain is the result of a p-transposable element insertion. No information about the phenotype is publicly available.
Fifteen other mutant alleles are known, but are less commonly researched.
See also
References
- ↑ "cyc cycle [Drosophila melanogaster (fruit fly)]". cyc cyle gene. National Center for Biotechnology Information, U.S. National Library of Medicine. Retrieved 10 April 2013.
- ↑ "Transcript: cyc-RA FBtr0074924". cyc-RA FBtr0074924. Ensembl. Retrieved 10 April 2013.
- 1 2 "Dmel/cyc". FlyBase Gene Report: Dmel/cyc. The Genetics Society of America.|accessdate=10 April 2013
- 1 2 3 4 5 6 7 Rutila JE, Suri V, Le M, So WV, Rosbash M, Hall JC (May 1998). "CYCLE is a second bHLH-PAS clock protein essential for circadian rhythmicity and transcription of Drosophila period and timeless". Cell. 93 (5): 805–14. doi:10.1016/S0092-8674(00)81441-5. PMID 9630224.
- ↑ Maurer C, Hung HC, Weber F (May 2009). "Cytoplasmic interaction with CYCLE promotes the post-translational processing of the circadian CLOCK protein". FEBS Lett. 583 (10): 1561–6. doi:10.1016/j.febslet.2009.04.013. PMID 19376119.
- ↑ Keene AC, Duboué ER, McDonald DM, Dus M, Suh GS, Waddell S, Blau J (July 2010). "Clock and cycle limit starvation-induced sleep loss in Drosophila". Curr. Biol. 20 (13): 1209–15. doi:10.1016/j.cub.2010.05.029. PMC 2929698
 . PMID 20541409.
. PMID 20541409. - 1 2 Shaw PJ, Tononi G, Greenspan RJ, Robinson DF (May 2002). "Stress response genes protect against lethal effects of sleep deprivation in Drosophila". Nature. 417 (6886): 287–91. doi:10.1038/417287a. PMID 12015603.
- ↑ Darlington TK, Wager-Smith K, Ceriani MF, Staknis D, Gekakis N, Steeves TD, Weitz CJ, Takahashi JS, Kay SA (June 1998). "Closing the circadian loop: CLOCK-induced transcription of its own inhibitors per and tim". Science. 280 (5369): 1599–603. doi:10.1126/science.280.5369.1599. PMID 9616122.
- ↑ Allada R, White NE, So WV, Hall JC, Rosbash M (May 1998). "A mutant Drosophila homolog of mammalian Clock disrupts circadian rhythms and transcription of period and timeless". Cell. 93 (5): 791–804. doi:10.1016/S0092-8674(00)81440-3. PMID 9630223.
- ↑ Rakshit K, Krishnan N, Guzik EM, Pyza E, Giebultowicz JM (February 2012). "Effects of aging on the molecular circadian oscillations in Drosophila". Chronobiol. Int. 29 (1): 5–14. doi:10.3109/07420528.2011.635237. PMC 3265550. PMID 22217096.
- ↑ Collins B, Kane EA, Reeves DC, Akabas MH, Blau J (May 2012). "Balance of activity between LN(v)s and glutamatergic dorsal clock neurons promotes robust circadian rhythms in Drosophila". Neuron. 74 (4): 706–18. doi:10.1016/j.neuron.2012.02.034. PMC 3361687. PMID 22632728.
- ↑ Linford NJ, Chan TP, Pletcher SD (2012). "Re-patterning sleep architecture in Drosophila through gustatory perception and nutritional quality". PLoS Genet. 8 (5): e1002668. doi:10.1371/journal.pgen.1002668. PMC 3342939. PMID 22570630.
- ↑ Wang H (May 2009). "Comparative genomic analysis of teleost fish bmal genes". Genetica. 136 (1): 149–61. doi:10.1007/s10709-008-9328-9. PMID 18850331.
- ↑ "ARNTL Gene". ARNTL Gene. National Center for Biotechnology Information, U.S. National Library of Medicine. Retrieved 9 April 2013.
- ↑ Park JH, Helfrich-Förster C, Lee G, Liu L, Rosbash M, Hall JC (March 2000). "Differential regulation of circadian pacemaker output by separate clock genes in Drosophila". Proc. Natl. Acad. Sci. U.S.A. 97 (7): 3608–13. doi:10.1073/pnas.070036197. PMC 16287. PMID 10725392.
- ↑ Tanoue S, Krishnan P, Krishnan B, Dryer SE, Hardin PE (April 2004). "Circadian clocks in antennal neurons are necessary and sufficient for olfaction rhythms in Drosophila". Curr. Biol. 14 (8): 638–49. doi:10.1016/j.cub.2004.04.009. PMID 15084278.