Lanosterol 14 alpha-demethylase
| Cytochrome P450, Family 51, Subfamily A, Polypeptide 1 | |
|---|---|
| Identifiers | |
| Symbol | CYP51A1 |
| Alt. symbols | CYP51, P45014DM |
| Entrez | 1595 |
| HUGO | 2649 |
| OMIM | 601637 |
| RefSeq | NM_000786 |
| UniProt | Q16850 |
| Other data | |
| EC number | 1.14.13.70 |
| Locus | Chr. 7 q21.2-21.3 |
Lanosterol 14 α-demethylase (or CYP51A1) is a cytochrome P450 enzyme that is involved in the conversion of lanosterol to 4,4-dimethylcholesta-8(9),14,24-trien-3β-ol.[1] The cytochrome P450 isoenzymes are a conserved group of proteins that serve as key players in the metabolism of organic substances and the biosynthesis of important steroids, lipids, and vitamins in eukaryotes.[2] As a member of this family, lanosterol 14 α-demethylase is responsible for an essential step in the biosynthesis of sterols. In particular, this protein catalyzes the removal of the C-14 α-methyl group from lanosterol (Lepesheva et al.). This demethylation step is regarded as the initial checkpoint in the transformation of lanosterol to other sterols that are widely used within the cell (Lepesheva et al.).
Although lanosterol 14 α-demethylase is present in a wide variety of organisms, this enzyme is studied primarily in the context of fungi, where it plays an essential role in mediating membrane permeability.[3] In fungi, CYP51 catalyzes the demethylation of lanosterol to create an important precursor that is eventually converted into ergosterol (Lepesheva et al.). This steroid then makes its way throughout the cell, where it alters the permeability and rigidity of plasma membranes much as cholesterol does in animals.[4] Because ergosterol constitutes a fundamental component of fungal membranes, many antifungal medications have been developed to inhibit 14 α-demethylase activity and prevent the production of this key compound (Becher et al.).
Evolution
The structural and functional properties of the cytochrome P450 superfamily have been subject to extensive diversification over the course of evolution (Becher et al.). Recent estimates indicate that there are currently 10 classes and 267 families of CYP proteins.[5] It is believed that 14 α-demethylase or CYP51 diverged early in the cytochrome's evolutionary history and has preserved its function ever since; namely, the removal of the 14 α-methyl group from sterol substrates (Becher et al.).
Although CYP51's mode of action has been well conserved, the protein's sequence varies considerably between biological kingdoms.[6] CYP51 sequence comparisons between kingdoms reveal only a 22-30% similarity in amino acid composition.[7]
Enzyme structure
.png)
Although the structure of 14 α-demethylase may vary substantially from one organism to the next, sequence alignment analysis reveals that there are six regions in the protein that are highly conserved in eukaryotes (Lepesheva et al.). These include residues in the B' helix, B'/C loop, C helix, I helix, K/β1-4 loop, and β-strand 1-4 that are responsible for forming the surface of the substrate binding cavity (Becher et al.). Homology modeling reveals that substrates migrate from the surface of the protein to the enzyme's buried active site through a channel that is formed in part by the A' alpha helix and the β4 loop.[8][9] Finally, the active site contains a heme prosthetic group in which the iron is tethered to a thiolate ligand on a conserved cysteine residue (Lepesheva et al.). This group also binds diatomic oxygen at the sixth coordination site, which is eventually incorporated onto the substrate (Lepesheva et al.).
Enzyme mechanism
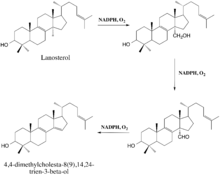
The enzyme-catalyzed demethylation of lanosterol is believed to occur in three steps, each of which requires one molecule of diatomic oxygen and one molecule of NADPH (or some other reducing equivalent).[10] During the first two steps, the 14α-methyl group undergoes typical cytochrome monooxygenation in which one oxygen atom is incorporated by the substrate and the other is reduced to water, resulting in the sterol's conversion to a carboxyalcohol and then a carboxyaldehyde (Lepesheva et al.). The aldehyde then departs as formic acid and a double bond is simultaneously introduced to yield the demethylated product (Lepesheva et al.).
Biological function
The biological role of this protein is also well-understood. The demethylated products of the CYP51 reaction are vital intermediates in pathways leading to the formation of cholesterol in humans, ergosterol in fungi, and other types of sterols in plants (Lepesheva et al.). These sterols localize to the plasma membrane of cells, where they play an important structural role in the regulation of membrane fluidity and permeability and also influence the activity of enzymes, ion channels, and other cell components that are embedded within (Daum et al.).[11][12] With the proliferation of immuno-suppressive diseases such as HIV/AIDS and cancer, patients have become increasingly vulnerable to opportunistic fungal infections (Richardson et al.). Seeking new means to treat such infections, drug researchers have begun targeting the 14 α-demethylase enzyme in fungi; destroying the fungal cell's ability to produce ergosterol causes a disruption of the plasma membrane, thereby resulting in cellular leakage and ultimately the death of the pathogen (DrugBank).
Azoles are currently the most popular class of antifungals used in both agricultural and medical settings (Becher et al.). These compounds bind as the sixth ligand to the heme group in CYP51, thereby altering the structure of the active site and acting as noncompetitive inhibitors.[13] The effectiveness of imidazoles and triazoles (common azole subclasses) as inhibitors of 14 α-demethylase have been confirmed through several experiments. Some studies test for changes in the production of important downstream ergosterol intermediates in the presence of these compounds.[14] Other studies employ spectrophotometry to quantify azole-CYP51 interactions (Becher et al.). Coordination of azoles to the prosthetic heme group in the enzyme's active site causes a characteristic shift in CYP51 absorbance, creating what is commonly referred to as a type II difference spectrum.[15][16]
Prolonged use of azoles as antifungals has resulted in the emergence of drug resistance among certain fungal strains (Becher et al.). Mutations in the coding region of CYP51 genes, overexpression of CYP51, and overexpression of membrane efflux transporters can all lead to resistance to these antifungals.[17][18][19][20] Consequently, the focus of azole research is beginning to shift towards identifying new ways to circumvent this major obstacle (Becher et al.).
See also
References
- ↑ "Metabocard for 4,4-Dimethylcholesta-8,14,24-trienol (HMDB01023)." Human Metabolome Database. Web. 25 Feb. 2014. <http://www.hmdb.ca/metabolites/HMDB01023>.
- ↑ Lepesheva, Galina I., and Michael R. Waterman. "Sterol 14α-Demethylase Cytochrome P450 (CYP51), a P450 in All Biological Kingdoms." Biochim Biophys Acta. 2008. 1770(3): 467-77.
- ↑ Daum G, Lees ND, Bard M, Dickson R. "Biochemistry, Cell Biology and Molecular Biology of Lipids of Saccharomyces cervisiae". Yeast. 1998. 14(16):1471-1510.
- ↑ Becher, Rayko, and Stefan G. R. Wirsel. "Fungal Cytochrome P450 Sterol 14α-demethylase (CYP51) and Azole Resistance in Plant and Human Pathogens." Applied Microbiology and Biotechnology. 2012. 95(4): 825-40.
- ↑ Hannemann F, Bichet A, Ewen KM, Bernhardt R. "Cytochrome P450 Systems—Biological Variations of Electron Transport Chains. Biochim Biophys Acta Gen Subj. 2007, 1770(3):330–344
- ↑ Lepesheva GI, Waterman MR. "CYP51—The Omnipotent P450." Mol Cell Endocrinol. 2004, 215:165–170
- ↑ Lepesheva GI, Waterman MR. "Structural Basis for Conservation in the CYP51 Family." Biochim Biophys Acta Proteins Proteom. 2011, 1814:88–93
- ↑ Hargrove TY, Wawrzak Z, Liu JL, Nes WD, Waterman MR, Lepesheva GI. "Substrate Preferences and Catalytic Parameters Determined by Structural Characteristics of Sterol 14alpha-demethylase (CYP51) from Leishmania infantum." J Biol Chem. 2011, 286:26838–26848.
- ↑ Podust LM, von Kries JP, Eddine AN, Kim Y et al. "Small-Molecule Scaffolds for CYP51 Inhibitors Identified by High-Throughput Screening and Defined by X-ray Crystallography." Antimicrob Agents Chemother. 2007, 51(11): 3915-23.
- ↑ Vanden Bossche H, Koymans L. "Cytochromes P450 in Fungi." Mycoses. 1998, 41:32–38
- ↑ Abe F, Usui K, Hiraki T. "Fluconazole Modulates Membrane Rigidity, Heterogeneity, and Water Penetration into the Plasma Membrane in Saccharomyces cerevisiae." Biochemistry. 2009, 48:8494–8504
- ↑ "Itraconazole (DB01167)." DrugBank. 25 Feb. 2014. <http://www.drugbank.ca/drugs/DB01167>.
- ↑ Mullins JGL, Parker JE, Cools HJ, Togawa RC, Lucas JA, et al. "Molecular Modeling of the Emergence of Azole Resistance in Mycosphaerella graminicola." PLoS ONE. 2011, 6(6): e20973. doi:10.1371/journal.pone.0020973
- ↑ Tuck SF, Patel H, Safi E, Robinson CH. "Lanosterol 14 Alpha-Demethylase (P45014DM): Effects of P45014DM Inhibitors on Sterol Biosynthesis Downstream of Lanosterol." J Lipid Res. 1991, 32(6): 893-902
- ↑ Vanden Bossche H, Marichal P, Gorrens J, Bellens D, Verhoeven H, Coene MC, Lauwers W, Janssen PAJ. "Interaction of Azole Derivatives with Cytochrome P-450 Isozymes in Yeast, Fungi, Plants and Mammalian-cells. Pestic Sci. 1987, 21:289–306
- ↑ Yoshida Y, Aoyama Y. "Interaction of Azole Antifungal Agents with Cytochrome P-45014DM Purified from Saccharomyces cerevisiae Microsomes." Biochem Pharmacol. 1987, 36:229–235
- ↑ Vanden Bossche H, Dromer F, Improvisi I, Lozano-Chiu M, Rex JH, Sanglard D. "Antifungal Drug Resistance in Pathogenic Fungi". Med Mycol. 1998, 36(1): 119–28.
- ↑ Leroux P, Albertini C, Gautier A, Gredt M, Walker AS. "Mutations in the CYP51 Gene Correlated with Changes in Sensitivity to Sterol 14Alpha-Demethylation Inhibitors in Field Isolates of Mycosphaerella graminicola." Pest Manag Sci. 2007, 63:688–698
- ↑ Sanglard D, Ischer F, Koymans L, Bille J. "Amino Acid Substitutions in the Cytochrome P-450 Lanosterol 14Alpha-Demethylase (CYP51A1) from Azole-resistant Candida albicans Clinical Isolates Contribute to Resistance to Azole Antifungal Agents. Antimicrob Agents Chemother. 1998, 42:241–253
- ↑ Cannon RD, Lamping E, Holmes AR, Niimi K, Baret PV, Keniya MV, Tanabe K, Niimi M, Goffeau A, Monk BC. "Efflux-mediated Antifungal Drug Resistance. Clin Microbiol Rev. 2009, 22:291– 321
External links
- cytochrome P-450 CYP51 at the US National Library of Medicine Medical Subject Headings (MeSH)