Loss of heterozygosity
Loss of heterozygosity (LOH) is a gross chromosomal event that results in loss of the entire gene and the surrounding chromosomal region.[1]
Most diploid cells, for example human somatic cells, contain two copies of the genome, one from each parent (chromosome pair); each copy contains approximately 3 billion bases (adenine (A), guanine (G), cytosine (C) or thymine (T)). For the majority of positions in the genome the base present is consistent between individuals, however a small percentage may contain different bases (usually one of two; for instance, ‘A’ or ‘G’) and these positions are called ‘single nucleotide polymorphisms’ or ‘SNPs’. When the genomic copies derived from each parent have different bases for these polymorphic regions (SNPs) the region is said to be heterozygous. Most of the chromosomes within somatic cells of individuals are paired, allowing for SNP locations to be potentially heterozygous. However, one parental copy of a region can sometimes be lost, which results in the region having just one copy. The single copy cannot be heterozygous at SNP locations and therefore the region shows loss of heterozygosity (LOH). Loss of heterozygosity due to loss of one parental copy in a region is also called hemizygosity in that region.
In cancer
The loss of heterozygosity is a common occurrence in cancer, where it indicates the absence of a functional tumor suppressor gene in the lost region. However, many people remain healthy with such a loss, because there still is one functional gene left on the other chromosome of the chromosome pair. However, the remaining copy of the tumor suppressor gene can be inactivated by a point mutation, leaving no tumor suppressor gene to protect the body. Loss of heterozygosity does not imply a reversal to the homozygous state.
Knudson two-hit hypothesis of tumorigenesis
- First Hit: The first hit is classically thought of as a point mutation that inactivates one copy of a tumor suppressor gene (TSG), such as Rb1. In hereditary cancer syndromes, individuals are born with the first hit. The individual does not develop cancer at this point because the remaining TSG allele on the other locus is still functioning normally.
- Second Hit: While the second hit is commonly assumed to be a deletion that results in loss of the remaining functioning TSG allele, the original published mechanism of RB1 LOH was mitotic recombination/gene conversion/copy-neutral LOH, not deletion. There is a critical difference between deletion and CN-LOH, as the latter mechanism cannot be detected by comparative genomic hybridization (CGH)-based gene copy number counting, and requires allelic genotyping. Either way, LOH leaves only non-functioning alleles of the TSG, and the individual goes on to develop cancer.
Copy-neutral LOH
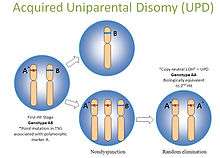
Copy-neutral LOH is thus called because no net change in the copy number occurs in the affected individual. Other names for copy-neutral LOH are acquired uniparental disomy (UPD) or gene conversion. In UPD, a person receives two copies of a chromosome, or part of a chromosome, from one parent and no copies from the other parent due to errors in meiosis I or meiosis II. This acquired homozygosity could lead to development of cancer if the individual inherited a non-functional allele of a tumor suppressor gene.
In tumor cells copy-neutral LOH can be biologically equivalent to the second hit in the Knudson hypothesis.[2] Acquired UPD is quite common in both hematologic and solid tumors, and is reported to constitute 20 to 80% of the LOH seen in human tumors.[3][4][5][6] Determination of virtual karyotypes using SNP-based arrays can provide genome-wide copy number and LOH status, including detection of copy-neutral LOH. Copy-neutral LOH cannot be detected by arrayCGH, FISH, or conventional cytogenetics. SNP-based arrays are preferred for virtual karyotyping of tumors and can be performed on fresh or paraffin-embedded tissues.

Retinoblastoma
The classical example of such a loss of protecting genes is hereditary retinoblastoma, in which one parent's contribution of the tumor suppressor Rb1 is flawed. Although most cells will have a functional second copy, chance loss of heterozygosity events in individual cells almost invariably lead to the development of this retinal cancer in the young child.
Breast Cancer and BRCA1/2
The genes BRCA1 and BRCA2 show loss of heterozygosity in samplings of tumors from patients who have germline mutations. BRCA1/2 are genes that produce proteins which regulate the DNA repair pathway by binding to Rad51.
Detection
Loss of heterozygosity can be identified in cancers by noting the presence of heterozygosity at a genetic locus in an organism's germline DNA, and the absence of heterozygosity at that locus in the cancer cells. This is often done using polymorphic markers, such as microsatellites or single-nucleotide polymorphisms, for which the two parents contributed different alleles. Genome-wide LOH status of fresh or paraffin embedded tissue samples can be assessed by virtual karyotyping using SNP arrays.
In asexual organisms
It has been proposed that LOH may limit the longevity of asexual organisms.[7][8] The minor allele in heterozygous areas of the genome is likely to have mild fitness consequences compared to de-novo mutations because selection has had time to remove deleterious alleles. When allelic gene conversion removes the major allele at these sites organisms are likely to experience a mild decline in fitness. Because LOH is much more common than de-novo mutation, and because the fitness consequences are closer to neutrality, this process should drive Muller's ratchet more quickly than de-novo mutations. While this process has received little experimental investigation, it is known that major signature of asexuality in metazoan genomes appears to be genome wide LOH, a sort of anti-meselson effect.
See also
- Microsatellite instability
- Tumor suppressor gene
- Virtual Karyotype
- Knudson hypothesis
- Deletion (genetics)
References
- ↑ [Association of the autoimmune diseases scleroderma with an immunologic response to cancer,] Christine G. Joseph et al., Science, 343:152 (10 January 2014)
- ↑ Mao X, Young BD, Lu YJ. The application of single-nucleotide polymorphism microarrays in cancer research. Curr Genomics. 2007 Jun;8(4):219–28.
- ↑ Gondek LP, Tiu R, O'Keefe CL, Sekeres MA, Theil KS, Maciejewski JP. Chromosomal lesions and uniparental disomy detected by SNP arrays in MDS, MDS/MPD, and MDS-derived AML. Blood. 2008 Feb 1;111(3):1534–42.
- ↑ Beroukhim R, Lin M, Park Y, Hao K, Zhao X, Garraway LA, et al. Inferring loss-of-heterozygosity from unpaired tumors using high-density oligonucleotide SNP arrays. PLoS Comput. Biol. 2006 May;2(5):e41.
- ↑ Ishikawa S, Komura D, Tsuji S, Nishimura K, Yamamoto S, Panda B, et al. Allelic dosage analysis with genotyping microarrays. Biochem Biophys Res Commun. 2005 Aug 12;333(4):1309–14.
- ↑ Lo KC, Bailey D, Burkhardt T, Gardina P, Turpaz Y, Cowell JK. Comprehensive analysis of loss of heterozygosity events in glioblastoma using the 100K SNP mapping arrays and comparison with copy number abnormalities defined by BAC array comparative genomic hybridization. Genes Chromosomes Cancer. 2008 Mar;47(3):221–37.
- ↑ Tucker AE, Ackerman MA, Eads BD, Xu S, Lynch M. Population-genomic insights into the evolutionary origin and fate of obligately asexual Daphnia pulex. PNAS. 2013; 110:15740.
- ↑ Archetti M. Recombination and loss of complementation: A more than two-fold cost for parthenogenesis. J Evol Biol 2004; 17(5):1084–1097.
External links
| Wikimedia Commons has media related to Loss of heterozygosity. |
- "Long-term study of the clinical significance of loss of heterozygosity in childhood acute lymphoblastic leukemia" – Leukemia
- "Loss of heterozygosity identifies genetic changes in chronic myeloid disorders, including myeloproliferative disorders, myelodysplastic syndromes and chronic myelomonocytic leukemia" – Modern Pathology
- "Mapping loss of heterozygosity in normal human breast cells from BRCA1/2 carriers" – BJC
- "Loss of Heterozygosity Studies on Chromosome 12q in Disseminated Superficial Actinic Porokeratosis: Lessons to be Learned" – Journal of Investigative Dermatology