Microbial rhodopsin
.jpg)
Microbial rhodopsin, also known as type-I rhodopsin, is a photochemically active membrane protein composed of seven transmembrane alpha-helices with a retinal chromophore. It includes light-driven proton pumps, ion pumps and ion channels, as well as light sensors. Microbial rhodopsins are found in all three domains of life: Archaea, Bacteria, and Eukaryota, as well as in viruses. It is rare in complex multicellular organisms.[2][3]
Nomenclature
Rhodopsin was originally a synonym for "visual purple", a visual pigment (light-sensitive molecule) found in the retinas of frogs and other vertebrates, used for dim-light vision, and usually found in rod cells. This is still the meaning of rhodopsin in the narrow sense. In the broad sense, rhodopsin refers to any molecule consisting of an opsin and a chromophore (generally a variant of retinal). The term bacterial rhodopsin originally referred to the first microbial rhodopsin discovered, known today as bacteriorhodopsin. Since then, other microbial rhodopsins have been discovered, rendering the term bacterial rhodopsin ambiguous.[4][5]
Table
Below is a list of some of the more well-known microbial rhodopsins and some of their properties.
| Function | Name | Abbr. | Ref. |
|---|---|---|---|
| proton pump (H+) | bacteriorhodopsin | BR | [6] |
| proton pump (H+) | proteorhodopsin | PR | [6] |
| proton pump (H+) | archaerhodopsin | Arch | [7] |
| proton pump (H+) | xanthorhodopsin | xR | [8] |
| cation channel (+) | channelrhodopsin | ChR | [9] |
| cation pump (Na+) | Krokinobacter eikastus rhodopsin 2 | KR2 | [10] |
| anion pump (Cl-) | halorhodopsin | HR | [6] |
| photosensor | sensory rhodopsin I | SR-I | [6] |
| photosensor | sensory rhodopsin II | SR-II | [6] |
| photosensor | Neurospora opsin I | NOP-I | [9][11] |
| light-activated enzyme | rhodopsin guanylyl cyclase | RhGC | [12] |
The Ion-Translocating Microbial Rhodopsin Family
The Ion-translocating Microbial Rhodopsin (MR) Family (TC# 3.E.1) is a member of the TOG Superfamily of secondary carriers. Members of the MR family catalyze light-driven ion translocation across microbial cytoplasmic membranes or serve as light receptors. Most proteins of the MR family are all of about the same size (250-350 amino acyl residues) and possess seven transmembrane helical spanners with their N-termini on the outside and their C-termini on the inside. There are 9 subfamilies in the MR family:[13]
(1) Bacteriorhodopsins pump protons out of the cell;
(2) Halorhodopsins pump chloride (and other anions such as bromide, iodide and nitrate) into the cell;
(3) Sensory rhodopsins, which normally function as receptors for phototactic behavior, are capable of pumping protons out of the cell if dissociated from their transducer proteins;
(4) the Fungal Chaparones are stress-induced proteins of ill-defined biochemical function, but this subfamily also includes a H+-pumping rhodopsin;[14]
(5) the bacterial rhodopsin, called Proteorhodopsin, is a light-driven proton pump that functions as does bacteriorhodopsins;
(6) the Neurospora crassa retinal-containing receptor serves as a photoreceptor (Neurospora ospin I);[15]
(7) the green algal light-gated proton channel, Channelrhodpsin-1;
(8) Sensory rhodopsins from cyanobacteria.
(9) Light-activated rhodopsin/guanylyl cyclase
A phylogenetic analysis of microbial rhodopsins and a detailed analysis of potential examples of horizontal gene transfer have been published.[16]
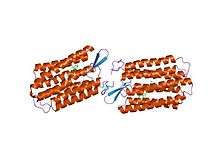
Structure
Among the high resolution structures for members of the MR Family are the archaeal proteins, bacteriorhodopsin (4QI1),[17] sensory rhodopsin II (3QAP),[18] halorhodopsin (3QBG) [19] as well as an Anabaena cyanobacterial sensory rhodopsin (2M3G) (TC# 3.E.1.1.6).[20]
A crystallographic structure of xanthorhodopsin at 1.9 Å resolution (3DDL) revealed a dual chromophore, the geometry of the carotenoid and the retinal.[21] The close approach of the 2 polyenes at their ring ends explains why the efficiency of the excited-state energy transfer is as high as approximately 45%, and the 46 degree angle between them suggests that the chromophore location is a compromise between optimal capture of light of all polarization angles and excited-state energy transfer. At 1.9 Å resolution, the structure revealed a light-driven proton pump with a dual chromophore. Ion-transporting rhodopsins of marine bacteria have been reviewed.[22][23]
There are several other crystal structures for members of the MR family available in RCSB. A list of currently available structures is also available in TCDB.
Function
The association of sensory rhodopsins with their transducer proteins appears to determine whether they function as transporters or receptors. Association of a sensory rhodopsin receptor with its transducer occurs via the transmembrane helical domains of the two interacting proteins. There are two sensory rhodopsins in any one halophilic archaeon, one (SRI) that responds positively to orange light but negatively to blue light, the other (SRII) that responds only negatively to blue light. Each transducer is specific for its cognate receptor. An x-ray structure of SRII complexed with its transducer (HtrII) at 1.94 Å resolution is available (1H2S).[24] Molecular and evolutionary aspects of the light-signal transduction by microbial sensory receptors have been reviewed.[23]
Homologues
Homologues include putative fungal chaperone proteins, a retinal-containing rhodopsin from Neurospora crassa,[25] a H+-pumping rhodopsin from Leptosphaeria maculans,[14] retinal-containing proton pumps isolated from marine bacteria,[26] a green light-activated photoreceptor in cyanobacteria that does not pump ions and interacts with a small (14 kDa) soluble transducer protein [20][27] and light-gated H+ channels from the green alga, Chlamydomonas reinhardtii.[28] The N. crassa NOP-1 protein exhibits a photocycle and conserved H+ translocation residues that suggest that this putative photoreceptor is a slow H+ pump.[14][29][30]
Most of the MR family homologues in yeast and fungi are of about the same size and topology as the archaeal proteins (283-344 amino acyl residues; 7 putative transmembrane α-helical segments), but they are heat shock- and toxic solvent-induced proteins of unknown biochemical function. They have been suggested to function as pmf-driven chaperones that fold extracellular proteins, but only indirect evidence supports this postulate.[15] The MR family is distantly related to the 7 TMS LCT family (TC# 2.A.43).[15]
Representative members of MR family can be found in the Transporter Classification Database.
Anabaena sensory rhodopsin
The Anabaena sensory rhodopsin exhibits light-induced interconversion between 13-cis and all trans states.[20] The ratio of its cis and trans chromophore forms depends on the wavelength of illumination, thus providing a mechanism for a single protein to signal the color of light, for example, to regulate color-sensitive processes such as chromatic adaptation in photosynthesis. Its cytoplasmic half channel, highly hydrophobic in the archaeal rhodopsins, contains numerous hydrophilic residues networked by water molecules, providing a connection from the photoactive site to the cytoplasmic surface believed to interact with the receptor's soluble 14-kilodalton transducer.
Bacteriorhodopsins and Halorhodopsins
Bacterio- and halorhodopsins pump 1 H+ and 1 Cl− per photon absorbed, respectively. Specific transport mechanisms and pathways have been proposed.[19][31][32] The mechanism involves:
(1) photo-isomerization of the retinal and its initial configurational changes,
(2) deprotonation of the retinal Schiff base and the coupled release of a proton to the extracellular membrane surface,
(3) the switch event that allows reprotonation of the Schiff base from the cytoplasmic side.
Six structural models describe the transformations of the retinal and its interaction with water 402, Asp85, and Asp212 in atomic detail, as well as the displacements of functional residues farther from the Schiff base. The changes provide rationales for how relaxation of the distorted retinal causes movements of water and protein atoms that result in vectorial proton transfers to and from the Schiff base.[31] Helix deformation is coupled to vectorial proton transport in the photocycle of bacteriorhodopsin.[33]
Most residues participating in the trimerization are not conserved in bacteriorhodopsin, a homologous protein capable of forming a trimeric structure in the absence of bacterioruberin. Despite a large alteration in the amino acid sequence, the shape of the intratrimer hydrophobic space filled by lipids is highly conserved between archaerhodopsin-2 and bacteriorhodopsin. Since a transmembrane helix facing this space undergoes a large conformational change during the proton pumping cycle, it is feasible that trimerization is an important strategy to capture special lipid components that are relevant to the protein activity.[34]
Marine Bacterial Rhodopsin
A marine bacterial rhodopsin has been reported to function as a proton pump. However, it also resembles sensory rhodopsin II of archaea as well as an Orf from the fungus Leptosphaeria maculans (AF290180). These proteins exhibit 20-30% identity with each other.
Channelrhodopsins
Channelrhodopsin-1 (ChR1) or channelopsin-1 (Chop1; Cop3; CSOA) of C. reinhardtii is closely related to the archaeal sensory rhodopsins. It has 712 aas with a signal peptide, followed by a short amphipathic region, and then a hydrophobic N-terminal domain with seven probable TMSs (residues 76-309) followed by a long hydrophilic C-terminal domain of about 400 residues. Part of the C-terminal hydrophilic domain is homologous to intersectin (EH and SH3 domain protein 1A) of animals (AAD30271).
Chop1 serves as a light-gated proton channel and mediates phototaxis and photophobic responses in green algae.[28] Based on this phenotype, Chop1 could be assigned to TC category #1.A, but because it belongs to a family in which well-characterized homologues catalyze active ion transport, it is assigned to the MR family. Expression of the chop1 gene, or a truncated form of that gene encoding only the hydrophobic core (residues 1-346 or 1-517) in frog oocytes in the presence of all-trans retinal produces a light-gated conductance that shows characteristics of a channel passively but selectively permeable to protons. This channel activity probably generates bioelectric currents.[28]
A homologue of ChR1 in C. reinhardtii is channelrhodopsin-2 (ChR2; Chop2; Cop4; CSOB). This protein is 57% identical, 10% similar to ChR1. It forms a cation-selective ion channel activated by light absorption. It transports both monovalent and divalent cations. It desensitizes to a small conductance in continuous light. Recovery from desensitization is accelerated by extracellular H+ and a negative membrane potential. It may be a photoreceptor for dark adapted cells.[35] A transient increase in hydration of transmembrane α-helices with a t(1/2) = 60 μs tallies with the onset of cation permeation. Aspartate 253 accepts the proton released by the Schiff base (t(1/2) = 10 μs), with the latter being reprotonated by aspartic acid 156 (t(1/2) = 2 ms). The internal proton acceptor and donor groups, corresponding to D212 and D115 in bacteriorhodopsin, are clearly different from other microbial rhodopsins, indicating that their spatial positions in the protein were relocated during evolution. E90 deprotonates exclusively in the nonconductive state. The observed proton transfer reactions and the protein conformational changes relate to the gating of the cation channel.[36]
Archaerhodopsin
Archaerhodopsin-2 (aR2), a retinal protein-carotenoid complex found in the claret membrane of Halorubrum sp. aus-2, functions as a light-driven proton pump. Trigonal and hexagonal crystals revealed that trimers are arranged on a honeycomb lattice.[34] In these crystals, the carotenoid bacterioruberin binds to crevices between the subunits of the trimer. Its polyene chain is inclined from the membrane normal by an angle of about 20 degrees and, on the cytoplasmic side, it is surrounded by helices AB and DE of neighboring subunits. This peculiar binding mode suggests that bacterioruberin plays a structural role for the trimerization of aR2. When compared with the aR2 structure in another crystal form containing no bacterioruberin, the proton release channel takes a more closed conformation in the P321 or P6(3) crystal; i.e., the native conformation of protein is stabilized in the trimeric protein-bacterioruberin complex.
Transport Reaction
The generalized transport reaction for bacterio- and sensory rhodopsins is:[13]
- H+ (in) + hν → H+ (out).
That for halorhodopsin is:
- Cl− (out) + hν → Cl− (in).
See also
- Bacteriorhodopsin
- Proteorhodopsin
- Opsin
- Protein-coated disc
References
- ↑ Oren, Aharon (2002). "Molecular ecology of extremely halophilic Archaea and Bacteria". FEMS Microbiology Ecology. 39 (1): 1–7. doi:10.1111/j.1574-6941.2002.tb00900.x. ISSN 0168-6496.
- ↑ Boeuf, Dominique; Audic, Stéphane; Brillet-Guéguen, Loraine; Caron, Christophe; Jeanthon, Christian (2015). "MicRhoDE: a curated database for the analysis of microbial rhodopsin diversity and evolution". Database. 2015: bav080. doi:10.1093/database/bav080. ISSN 1758-0463.
- ↑ Yawo, Hiromu; Kandori, Hideki; Koizumi, Amane (5 June 2015). Optogenetics: Light-Sensing Proteins and Their Applications. Springer. pp. 3–4. ISBN 978-4-431-55516-2. Retrieved 30 September 2015.
- ↑ "rhodopsin, n.". OED Online. Oxford University Press. 19 December 2012.
- ↑ Mason, Peggy (26 May 2011). Medical Neurobiology. OUP USA. p. 375. ISBN 978-0-19-533997-0. Retrieved 21 September 2015.
- 1 2 3 4 5 Yoshizawa, S.; Kumagai, Y.; Kim, H.; Ogura, Y.; Hayashi, T.; Iwasaki, W.; DeLong, E. F.; Kogure, K. (2014). "Functional characterization of flavobacteria rhodopsins reveals a unique class of light-driven chloride pump in bacteria". Proceedings of the National Academy of Sciences. 111 (18): 6732–6737. doi:10.1073/pnas.1403051111. ISSN 0027-8424.
- ↑ Zhang, Feng; Vierock, Johannes; Yizhar, Ofer; Fenno, Lief E.; Tsunoda, Satoshi; Kianianmomeni, Arash; Prigge, Matthias; Berndt, Andre; Cushman, John; Polle, Jürgen; Magnuson, Jon; Hegemann, Peter; Deisseroth, Karl (2011). "The Microbial Opsin Family of Optogenetic Tools". Cell. 147 (7): 1446–1457. doi:10.1016/j.cell.2011.12.004. ISSN 0092-8674.
- ↑ Sudo, Y.; Ihara, K.; Kobayashi, S.; Suzuki, D.; Irieda, H.; Kikukawa, T.; Kandori, H.; Homma, M. (2010). "A Microbial Rhodopsin with a Unique Retinal Composition Shows Both Sensory Rhodopsin II and Bacteriorhodopsin-like Properties". Journal of Biological Chemistry. 286 (8): 5967–5976. doi:10.1074/jbc.M110.190058. ISSN 0021-9258.
- 1 2 Heintzen, Christian (2012). "Plant and fungal photopigments". Wiley Interdisciplinary Reviews: Membrane Transport and Signaling. 1 (4): 411–432. doi:10.1002/wmts.36. ISSN 2190-460X.
- ↑ Kato, Hideaki E.; Inoue, Keiichi; Abe-Yoshizumi, Rei; Kato, Yoshitaka (2015). "Structural basis for Na+ transport mechanism by a light-driven Na+ pump". Nature. 521 (7550): 48–53. doi:10.1038/nature14322. ISSN 0028-0836.
- ↑ Olmedo, María; Ruger-Herreros, Carmen; Luque, Eva M.; Corrochano, Luis M. (2013). "Regulation of transcription by light in Neurospora crassa: A model for fungal photobiology?" (PDF). Fungal Biology Reviews. 27 (1): 10–18. doi:10.1016/j.fbr.2013.02.004. ISSN 1749-4613.
- ↑ Scheib, U; Stehfest, K; Gee, CE; Koerschen, HG; Fudim, R; Oertner, TO; Hegemann, P (2015). "The rhodopsin-guanylyl cyclase of the aquatic fungus Blastocladiella emersonii enables fast optical control of cGMP signaling". Science Signaling. 8 (389): rs8. doi:10.1126/scisignal.aab0611.
- 1 2 Saier, MH Jr. "3.E.1 The Ion-translocating Microbial Rhodopsin (MR) Family". Transporter Classification Database. Saier Lab Bioinformatics Group / SDSC.
- 1 2 3 Waschuk, Stephen A.; Bezerra, Arandi G.; Shi, Lichi; Brown, Leonid S. (2005-05-10). "Leptosphaeria rhodopsin: bacteriorhodopsin-like proton pump from a eukaryote". Proceedings of the National Academy of Sciences of the United States of America. 102 (19): 6879–6883. doi:10.1073/pnas.0409659102. ISSN 0027-8424. PMC 1100770
 . PMID 15860584.
. PMID 15860584. - 1 2 3 Zhai, Y.; Heijne, W. H.; Smith, D. W.; Saier, M. H. (2001-04-02). "Homologues of archaeal rhodopsins in plants, animals and fungi: structural and functional predications for a putative fungal chaperone protein". Biochimica et Biophysica Acta. 1511 (2): 206–223. doi:10.1016/s0005-2736(00)00389-8. ISSN 0006-3002. PMID 11286964.
- ↑ Sharma, Adrian K.; Spudich, John L.; Doolittle, W. Ford (2006-11-01). "Microbial rhodopsins: functional versatility and genetic mobility". Trends in Microbiology. 14 (11): 463–469. doi:10.1016/j.tim.2006.09.006. ISSN 0966-842X. PMID 17008099.
- ↑ Luecke, H.; Schobert, B.; Richter, H. T.; Cartailler, J. P.; Lanyi, J. K. (1999-10-08). "Structural changes in bacteriorhodopsin during ion transport at 2 angstrom resolution". Science (New York, N.Y.). 286 (5438): 255–261. doi:10.1126/science.286.5438.255. ISSN 0036-8075. PMID 10514362.
- ↑ Royant, A.; Nollert, P.; Edman, K.; Neutze, R.; Landau, E. M.; Pebay-Peyroula, E.; Navarro, J. (2001-08-28). "X-ray structure of sensory rhodopsin II at 2.1-A resolution". Proceedings of the National Academy of Sciences of the United States of America. 98 (18): 10131–10136. doi:10.1073/pnas.181203898. ISSN 0027-8424. PMC 56927. PMID 11504917.
- 1 2 Kolbe, M.; Besir, H.; Essen, L. O.; Oesterhelt, D. (2000-05-26). "Structure of the light-driven chloride pump halorhodopsin at 1.8 A resolution". Science (New York, N.Y.). 288 (5470): 1390–1396. doi:10.1126/science.288.5470.1390. ISSN 0036-8075. PMID 10827943.
- 1 2 3 Vogeley, Lutz; Sineshchekov, Oleg A.; Trivedi, Vishwa D.; Sasaki, Jun; Spudich, John L.; Luecke, Hartmut (2004-11-19). "Anabaena sensory rhodopsin: a photochromic color sensor at 2.0 A". Science (New York, N.Y.). 306 (5700): 1390–1393. doi:10.1126/science.1103943. ISSN 1095-9203. PMID 15459346.
- ↑ Luecke, Hartmut; Schobert, Brigitte; Stagno, Jason; Imasheva, Eleonora S.; Wang, Jennifer M.; Balashov, Sergei P.; Lanyi, Janos K. (2008-10-28). "Crystallographic structure of xanthorhodopsin, the light-driven proton pump with a dual chromophore". Proceedings of the National Academy of Sciences of the United States of America. 105 (43): 16561–16565. doi:10.1073/pnas.0807162105. ISSN 1091-6490. PMC 2575459. PMID 18922772.
- ↑ Inoue, Keiichi; Kato, Yoshitaka; Kandori, Hideki (2015-02-01). "Light-driven ion-translocating rhodopsins in marine bacteria". Trends in Microbiology. 23 (2): 91–98. doi:10.1016/j.tim.2014.10.009. ISSN 1878-4380. PMID 25432080.
- 1 2 Inoue, Keiichi; Tsukamoto, Takashi; Sudo, Yuki (2014-05-01). "Molecular and evolutionary aspects of microbial sensory rhodopsins". Biochimica et Biophysica Acta. 1837 (5): 562–577. doi:10.1016/j.bbabio.2013.05.005. ISSN 0006-3002. PMID 23732219.
- ↑ Gordeliy, Valentin I.; Labahn, Jörg; Moukhametzianov, Rouslan; Efremov, Rouslan; Granzin, Joachim; Schlesinger, Ramona; Büldt, Georg; Savopol, Tudor; Scheidig, Axel J. (2002-10-03). "Molecular basis of transmembrane signalling by sensory rhodopsin II-transducer complex". Nature. 419 (6906): 484–487. doi:10.1038/nature01109. ISSN 0028-0836. PMID 12368857.
- ↑ Maturana, A.; Arnaudeau, S.; Ryser, S.; Banfi, B.; Hossle, J. P.; Schlegel, W.; Krause, K. H.; Demaurex, N. (2001-08-10). "Heme histidine ligands within gp91(phox) modulate proton conduction by the phagocyte NADPH oxidase". The Journal of Biological Chemistry. 276 (32): 30277–30284. doi:10.1074/jbc.M010438200. ISSN 0021-9258. PMID 11389135.
- ↑ Béjà, O.; Aravind, L.; Koonin, E. V.; Suzuki, M. T.; Hadd, A.; Nguyen, L. P.; Jovanovich, S. B.; Gates, C. M.; Feldman, R. A. (2000-09-15). "Bacterial rhodopsin: evidence for a new type of phototrophy in the sea". Science (New York, N.Y.). 289 (5486): 1902–1906. doi:10.1126/science.289.5486.1902. ISSN 0036-8075. PMID 10988064.
- ↑ Jung, Kwang-Hwan; Trivedi, Vishwa D.; Spudich, John L. (2003-03-01). "Demonstration of a sensory rhodopsin in eubacteria". Molecular Microbiology. 47 (6): 1513–1522. doi:10.1046/j.1365-2958.2003.03395.x. ISSN 0950-382X. PMID 12622809.
- 1 2 3 Nagel, Georg; Ollig, Doris; Fuhrmann, Markus; Kateriya, Suneel; Musti, Anna Maria; Bamberg, Ernst; Hegemann, Peter (2002-06-28). "Channelrhodopsin-1: a light-gated proton channel in green algae". Science (New York, N.Y.). 296 (5577): 2395–2398. doi:10.1126/science.1072068. ISSN 1095-9203. PMID 12089443.
- ↑ Brown, L. S.; Dioumaev, A. K.; Lanyi, J. K.; Spudich, E. N.; Spudich, J. L. (2001-08-31). "Photochemical reaction cycle and proton transfers in Neurospora rhodopsin". The Journal of Biological Chemistry. 276 (35): 32495–32505. doi:10.1074/jbc.M102652200. ISSN 0021-9258. PMID 11435422.
- ↑ Brown, Leonid S. (2004-06-01). "Fungal rhodopsins and opsin-related proteins: eukaryotic homologues of bacteriorhodopsin with unknown functions". Photochemical & Photobiological Sciences: Official Journal of the European Photochemistry Association and the European Society for Photobiology. 3 (6): 555–565. doi:10.1039/b315527g. ISSN 1474-905X. PMID 15170485.
- 1 2 Lanyi, Janos K.; Schobert, Brigitte (2003-04-25). "Mechanism of proton transport in bacteriorhodopsin from crystallographic structures of the K, L, M1, M2, and M2' intermediates of the photocycle". Journal of Molecular Biology. 328 (2): 439–450. doi:10.1016/s0022-2836(03)00263-8. ISSN 0022-2836. PMID 12691752.
- ↑ Schobert, Brigitte; Brown, Leonid S.; Lanyi, Janos K. (2003-07-11). "Crystallographic structures of the M and N intermediates of bacteriorhodopsin: assembly of a hydrogen-bonded chain of water molecules between Asp-96 and the retinal Schiff base". Journal of Molecular Biology. 330 (3): 553–570. doi:10.1016/s0022-2836(03)00576-x. ISSN 0022-2836. PMID 12842471.
- ↑ Royant, A.; Edman, K.; Ursby, T.; Pebay-Peyroula, E.; Landau, E. M.; Neutze, R. (2000-08-10). "Helix deformation is coupled to vectorial proton transport in the photocycle of bacteriorhodopsin". Nature. 406 (6796): 645–648. doi:10.1038/35020599. ISSN 0028-0836. PMID 10949307.
- 1 2 Yoshimura, Keiko; Kouyama, Tsutomu (2008-02-01). "Structural role of bacterioruberin in the trimeric structure of archaerhodopsin-2". Journal of Molecular Biology. 375 (5): 1267–1281. doi:10.1016/j.jmb.2007.11.039. ISSN 1089-8638. PMID 18082767.
- ↑ Nagel, Georg; Szellas, Tanjef; Huhn, Wolfram; Kateriya, Suneel; Adeishvili, Nona; Berthold, Peter; Ollig, Doris; Hegemann, Peter; Bamberg, Ernst (2003-11-25). "Channelrhodopsin-2, a directly light-gated cation-selective membrane channel". Proceedings of the National Academy of Sciences of the United States of America. 100 (24): 13940–13945. doi:10.1073/pnas.1936192100. ISSN 0027-8424. PMC 283525. PMID 14615590.
- ↑ Lórenz-Fonfría, Víctor A.; Resler, Tom; Krause, Nils; Nack, Melanie; Gossing, Michael; Fischer von Mollard, Gabriele; Bamann, Christian; Bamberg, Ernst; Schlesinger, Ramona (2013-04-02). "Transient protonation changes in channelrhodopsin-2 and their relevance to channel gating". Proceedings of the National Academy of Sciences of the United States of America. 110 (14): E1273–1281. doi:10.1073/pnas.1219502110. ISSN 1091-6490. PMC 3619329. PMID 23509282.