PI3K/AKT/mTOR pathway
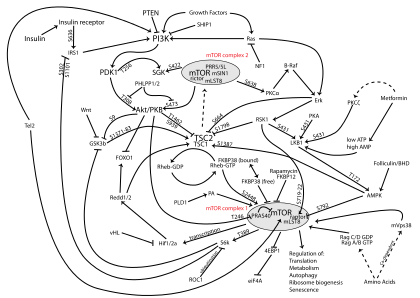
The PI3K/AKT/mTOR pathway is an intracellular signaling pathway important in regulating the cell cycle. Therefore, it is directly related to cellular quiescence, proliferation, cancer, and longevity. PI3K activation phosphorylates and activates AKT, localizing it in the plasma membrane.[1] AKT can have a number of downstream effects such as activating CREB,[2] inhibiting p27,[3] localizing FOXO in the cytoplasm,[3] activating PtdIns-3ps,[4] and activating mTOR [3] which can affect transcription of p70 or 4EBP1.[3] There are many known factors that enhance the PI3K/AKT pathway including EGF,[5] shh,[6] IGF-1,[7] insulin,[3] and CaM.[8] The pathway is antagonized by various factors including PTEN,[9] GSK3B,[10] and HB9.[11] In many cancers, this pathway is overactive, thus reducing apoptosis and allowing proliferation. This pathway is necessary, however, to promote growth and proliferation over differentiation of adult stem cells, neural stem cells specifically.[12] It is the difficulty in finding an appropriate amount of proliferation versus differentiation that researchers are trying to determine in order to utilize this balance in the development of various therapies.[13] Additionally, this pathway has been found to be a necessary component in neural long term potentiation.[14][15]
Proliferation of neural stem cells
Response to glucose
Neural stem cells (NSCs) in the brain must find a balance between maintaining their multipotency by self renewing and proliferating as opposed to differentiating and becoming quiescent. The PI3K/AKT pathway is crucial in this decision making process. NSCs are able to sense and respond to changes in the brain or throughout the organism. When glucose levels are high, insulin and therefore IGF is produced.[3] This signaling activates the PI3K/AKT pathway which works to promote proliferation. In this way, when there is high glucose and abundant energy in the organism, the PI3K/AKT pathway is activated and NSCs tend to proliferate. When there are low amounts of available energy, the PI3K/AKT pathway is less active and cells adopt a quiescent state. This occurs, in part, when AKT phosphorylates FOXO, keeping FOXO in the cytoplasm.[3] FOXO, when dephosphorylated, can enter the nucleus and work as a transcription factor to promote the expression of various tumor suppressors such as p27 and p21.[3] These tumor suppressors push the NSC to enter quiescence. FOXO knockouts lose the ability for cells to enter a quiescent state as well as cells losing their neural stem cell character, possibly entering a cancer like state.[3]
PTEN
The PI3K/AKT pathway has a natural inhibitor called PTEN whose function is to limit proliferation in cells, helping to prevent cancer. Knocking out PTEN has been shown to increase the mass of the brain because of the unregulated proliferation that occurs.[3] PTEN works by dephosphorylating PIP3 to PIP2 which limits AKTs ability to bind to the membrane, decreasing its activity. PTEN deficiencies can be compensated downstream to rescue differentiation or quiescence. Knocking out PTEN is not as serious as knocking out FOXO for this reason.[3]
CREB
The cAMP response element CREB is closely related to the cell decision to proliferate or not. Cells that are forced to overexpress AKT increase the amount of CREB and proliferation compared to wild type cells. These cells also express less glial and neural cell markers such as GFAP or β-tubulin.[16] This is because CREB is a transcription factor that influences the transcription of cyclin A which promotes proliferation.[17] For example, adult hippocampal neural progenitor cells need abeyance as stem cells to differentiate later. This is regulated by Shh. Shh works through a slow protein synthesis dependence, which stimulates other cascades that work synergistically with the PI3K/AKT pathway to induce proliferation. Then, the other pathway can be turned off and the effects of the PI3K/AKT pathway become insufficient in stopping differentiation.[18] The specifics of this pathway are unknown.
Therapies
Neural stem cells
The type of growth factor signaling can effect whether or not NSCs differentiate into motor neurons or not. Priming a media with FGF2 lowers the activity of the PI3K/AKT pathway, which activates GSK3β. This increases expression of HB9.[19] Directly inhibiting PI3K in NSCs leads to a population of cells that are purely HB9+ and differentiate at an elevated efficiency into motor neurons. Grafting these cells into different parts of rats generates motor neurons regardless of the transplanted cells' microenvironment.[20] Following injury, neural stem cells enter a repair phase and express high levels of PI3K to enhance proliferation. This is better for survival of the neurons as a whole but is at the expense of generating motor neurons. Therefore, it can be difficult for injured motor neurons to recover their ability.[21] It is the purpose of modern research to generate neural stem cells that can proliferate but still differentiate into motor neurons. Lowering the effect of the PI3K pathway and increasing the effect of GSK3β and HB9 in NSCs is a potential way of generating these cells.[22]
Bisperoxovanadium eg for neuroprotection after CNS injury
PTEN is a natural inhibitor of the PI3K/AKT pathway. Bisperoxovanadium is a specific inhibitor of PTEN's phosphatase activity[23] and has a definite half life. Therefore, by administering this PTEN inhibitor, one can temporarily and safely effect the PI3K/AKT pathway to influence cell migration,[24] survival [25] and proliferation.[26] Too much inhibition leads to unregulated cell cycle progression and tumorigenesis. However, enough PTEN inhibition promotes neuroprotection after CNS injury [27] and leads to an enhanced recovery to the CNS via axonal outgrowth.[28] Axons need to undergo outgrowth so that they can travel and connect to their targets, but lack the intrinsic capability to do so alone. Amplifying the PI3K/AKT pathway increases this neural outgrowth. It is the purpose of modern research to determine appropriate treatment concentrations of bisperoxovanadium to stimulate axonal outgrowth but not cause cancer.[29]
Long-term potentiation
In order for long-term potentiation (LTP) to occur, there must be stimulation of NMDA receptors, which causes AMPA receptors to be inserted postsynaptically. PI3K binds to AMPA receptors in a conserved region to orient the receptors in the membrane, specifically at the GluR subunit.[30] PI3K activity increases in response to calcium ions and CaM. Additionally, AKT localizes PtdIns-3Ps in the post synapse, which recruits docking proteins such as tSNARE and Vam7. This directly leads to the docking of AMPA in the post synapse.[31] mTOR activated p7056K and inactivated 4EBP1 which changes gene expression to allow LTP to occur.[32] Long-term fear conditioning training was affected in rats but there was no effect in short term conditioning. Specifically, amygdala fear conditioning was lost. This is a type of trace conditioning which is a form of learning that requires association of a conditioned stimulus with an unconditioned stimulus. This effect was lost in PI3K knockdowns and increased in PI3K overexpressions.[33]
See also
References
- ↑ King, D; Yeomanson, D; Bryant, HE (24 March 2015). "PI3King the Lock: Targeting the PI3K/Akt/mTOR Pathway as a Novel Therapeutic Strategy in Neuroblastoma". Journal of pediatric hematology/oncology. 37 (4): 245–51. doi:10.1097/MPH.0000000000000329. PMID 25811750.
- ↑ Peltier, J; O'Neill, A; Schaffer, D. V. (2007). "PI3K/Akt and CREB regulate adult neural hippocampal progenitor proliferation and differentiation". Developmental Neurobiology. 67 (10): 1348–61. doi:10.1002/dneu.20506. PMID 17638387.
- 1 2 3 4 5 6 7 8 9 10 11 Rafalski, V. A.; Brunet, A (2011). "Energy metabolism in adult neural stem cell fate". Progress in Neurobiology. 93 (2): 182–203. doi:10.1016/j.pneurobio.2010.10.007. PMID 21056618.
- ↑ Man, H. Y.; Wang, Q; Lu, W. Y.; Ju, W; Ahmadian, G; Liu, L; d'Souza, S; Wong, T. P.; Taghibiglou, C; Lu, J; Becker, L. E.; Pei, L; Liu, F; Wymann, M. P.; MacDonald, J. F.; Wang, Y. T. (2003). "Activation of PI3-kinase is required for AMPA receptor insertion during LTP of mEPSCs in cultured hippocampal neurons". Neuron. 38 (4): 611–24. doi:10.1016/s0896-6273(03)00228-9. PMID 12765612.
- ↑ Ojeda, L; Gao, J; Hooten, K. G.; Wang, E; Thonhoff, J. R.; Dunn, T. J.; Gao, T; Wu, P (2011). "Critical role of PI3K/Akt/GSK3β in motoneuron specification from human neural stem cells in response to FGF2 and EGF". PLoS ONE. 6 (8): e23414. doi:10.1371/journal.pone.0023414. PMC 3160859
 . PMID 21887250.
. PMID 21887250. - ↑ Peltier, J; O'Neill, A; Schaffer, D. V. (2007). "PI3K/Akt and CREB regulate adult neural hippocampal progenitor proliferation and differentiation". Developmental Neurobiology. 67 (10): 1348–61. doi:10.1002/dneu.20506. PMID 17638387.
- ↑ Peltier, J; O'Neill, A; Schaffer, D. V. (2007). "PI3K/Akt and CREB regulate adult neural hippocampal progenitor proliferation and differentiation". Developmental Neurobiology. 67 (10): 1348–61. doi:10.1002/dneu.20506. PMID 17638387.
- ↑ Man, H. Y.; Wang, Q; Lu, W. Y.; Ju, W; Ahmadian, G; Liu, L; d'Souza, S; Wong, T. P.; Taghibiglou, C; Lu, J; Becker, L. E.; Pei, L; Liu, F; Wymann, M. P.; MacDonald, J. F.; Wang, Y. T. (2003). "Activation of PI3-kinase is required for AMPA receptor insertion during LTP of mEPSCs in cultured hippocampal neurons". Neuron. 38 (4): 611–24. doi:10.1016/s0896-6273(03)00228-9. PMID 12765612.
- ↑ Wyatt, L. A.; Filbin, M. T.; Keirstead, H. S. (2014). "PTEN inhibition enhances neurite outgrowth in human embryonic stem cell-derived neuronal progenitor cells". Journal of Comparative Neurology. 522 (12): 2741–55. doi:10.1002/cne.23580. PMID 24610700.
- ↑ Peltier, J; O'Neill, A; Schaffer, D. V. (2007). "PI3K/Akt and CREB regulate adult neural hippocampal progenitor proliferation and differentiation". Developmental Neurobiology. 67 (10): 1348–61. doi:10.1002/dneu.20506. PMID 17638387.
- ↑ Ojeda, L; Gao, J; Hooten, K. G.; Wang, E; Thonhoff, J. R.; Dunn, T. J.; Gao, T; Wu, P (2011). "Critical role of PI3K/Akt/GSK3β in motoneuron specification from human neural stem cells in response to FGF2 and EGF". PLoS ONE. 6 (8): e23414. doi:10.1371/journal.pone.0023414. PMC 3160859. PMID 21887250.
- ↑ Peltier, J; O'Neill, A; Schaffer, D. V. (2007). "PI3K/Akt and CREB regulate adult neural hippocampal progenitor proliferation and differentiation". Developmental Neurobiology. 67 (10): 1348–61. doi:10.1002/dneu.20506. PMID 17638387.
- ↑ Peltier, J; O'Neill, A; Schaffer, D. V. (2007). "PI3K/Akt and CREB regulate adult neural hippocampal progenitor proliferation and differentiation". Developmental Neurobiology. 67 (10): 1348–61. doi:10.1002/dneu.20506. PMID 17638387.
- ↑ Man, H. Y.; Wang, Q; Lu, W. Y.; Ju, W; Ahmadian, G; Liu, L; d'Souza, S; Wong, T. P.; Taghibiglou, C; Lu, J; Becker, L. E.; Pei, L; Liu, F; Wymann, M. P.; MacDonald, J. F.; Wang, Y. T. (2003). "Activation of PI3-kinase is required for AMPA receptor insertion during LTP of mEPSCs in cultured hippocampal neurons". Neuron. 38 (4): 611–24. doi:10.1016/s0896-6273(03)00228-9. PMID 12765612.
- ↑ Sui, L; Wang, J; Li, B. M. (2008). "Role of the phosphoinositide 3-kinase-Akt-mammalian target of the rapamycin signaling pathway in long-term potentiation and trace fear conditioning memory in rat medial prefrontal cortex". Learning & Memory. 15 (10): 762–76. doi:10.1101/lm.1067808. PMID 18832563.
- ↑ Peltier, J; O'Neill, A; Schaffer, D. V. (2007). "PI3K/Akt and CREB regulate adult neural hippocampal progenitor proliferation and differentiation". Developmental Neurobiology. 67 (10): 1348–61. doi:10.1002/dneu.20506. PMID 17638387.
- ↑ Peltier, J; O'Neill, A; Schaffer, D. V. (2007). "PI3K/Akt and CREB regulate adult neural hippocampal progenitor proliferation and differentiation". Developmental Neurobiology. 67 (10): 1348–61. doi:10.1002/dneu.20506. PMID 17638387.
- ↑ Peltier, J; O'Neill, A; Schaffer, D. V. (2007). "PI3K/Akt and CREB regulate adult neural hippocampal progenitor proliferation and differentiation". Developmental Neurobiology. 67 (10): 1348–61. doi:10.1002/dneu.20506. PMID 17638387.
- ↑ Ojeda, L; Gao, J; Hooten, K. G.; Wang, E; Thonhoff, J. R.; Dunn, T. J.; Gao, T; Wu, P (2011). "Critical role of PI3K/Akt/GSK3β in motoneuron specification from human neural stem cells in response to FGF2 and EGF". PLoS ONE. 6 (8): e23414. doi:10.1371/journal.pone.0023414. PMC 3160859. PMID 21887250.
- ↑ Ojeda, L; Gao, J; Hooten, K. G.; Wang, E; Thonhoff, J. R.; Dunn, T. J.; Gao, T; Wu, P (2011). "Critical role of PI3K/Akt/GSK3β in motoneuron specification from human neural stem cells in response to FGF2 and EGF". PLoS ONE. 6 (8): e23414. doi:10.1371/journal.pone.0023414. PMC 3160859. PMID 21887250.
- ↑ Ojeda, L; Gao, J; Hooten, K. G.; Wang, E; Thonhoff, J. R.; Dunn, T. J.; Gao, T; Wu, P (2011). "Critical role of PI3K/Akt/GSK3β in motoneuron specification from human neural stem cells in response to FGF2 and EGF". PLoS ONE. 6 (8): e23414. doi:10.1371/journal.pone.0023414. PMC 3160859. PMID 21887250.
- ↑ Ojeda, L; Gao, J; Hooten, K. G.; Wang, E; Thonhoff, J. R.; Dunn, T. J.; Gao, T; Wu, P (2011). "Critical role of PI3K/Akt/GSK3β in motoneuron specification from human neural stem cells in response to FGF2 and EGF". PLoS ONE. 6 (8): e23414. doi:10.1371/journal.pone.0023414. PMC 3160859. PMID 21887250.
- ↑ Schmid, A. C.; Byrne, R. D.; Vilar, R; Woscholski, R (2004). "Bisperoxovanadium compounds are potent PTEN inhibitors". FEBS Letters. 566 (1–3): 35–8. doi:10.1016/j.febslet.2004.03.102. PMID 15147864.
- ↑ Mihai, C; Bao, S; Lai, J. P.; Ghadiali, S. N.; Knoell, D. L. (2012). "PTEN inhibition improves wound healing in lung epithelia through changes in cellular mechanics that enhance migration". AJP: Lung Cellular and Molecular Physiology. 302 (3): L287–99. doi:10.1152/ajplung.00037.2011. PMC 3289272. PMID 22037358.
- ↑ Lai, J. P.; Dalton, J. T.; Knoell, D. L. (2007). "Phosphatase and tensin homologue deleted on chromosome ten (PTEN) as a molecular target in lung epithelial wound repair". British Journal of Pharmacology. 152 (8): 1172–84. doi:10.1038/sj.bjp.0707501. PMC 2189995. PMID 17922022.
- ↑ Wyatt, L. A.; Filbin, M. T.; Keirstead, H. S. (2014). "PTEN inhibition enhances neurite outgrowth in human embryonic stem cell-derived neuronal progenitor cells". Journal of Comparative Neurology. 522 (12): 2741–55. doi:10.1002/cne.23580. PMID 24610700.
- ↑ Walker, C. L.; Walker, M. J.; Liu, N. K.; Risberg, E. C.; Gao, X; Chen, J; Xu, X. M. (2012). "Systemic bisperoxovanadium activates Akt/mTOR, reduces autophagy, and enhances recovery following cervical spinal cord injury". PLoS ONE. 7 (1): e30012. doi:10.1371/journal.pone.0030012. PMC 3254642. PMID 22253859.
- ↑ Wyatt, L. A.; Filbin, M. T.; Keirstead, H. S. (2014). "PTEN inhibition enhances neurite outgrowth in human embryonic stem cell-derived neuronal progenitor cells". Journal of Comparative Neurology. 522 (12): 2741–55. doi:10.1002/cne.23580. PMID 24610700.
- ↑ Wyatt, L. A.; Filbin, M. T.; Keirstead, H. S. (2014). "PTEN inhibition enhances neurite outgrowth in human embryonic stem cell-derived neuronal progenitor cells". Journal of Comparative Neurology. 522 (12): 2741–55. doi:10.1002/cne.23580. PMID 24610700.
- ↑ Man, H. Y.; Wang, Q; Lu, W. Y.; Ju, W; Ahmadian, G; Liu, L; d'Souza, S; Wong, T. P.; Taghibiglou, C; Lu, J; Becker, L. E.; Pei, L; Liu, F; Wymann, M. P.; MacDonald, J. F.; Wang, Y. T. (2003). "Activation of PI3-kinase is required for AMPA receptor insertion during LTP of mEPSCs in cultured hippocampal neurons". Neuron. 38 (4): 611–24. doi:10.1016/s0896-6273(03)00228-9. PMID 12765612.
- ↑ Man, H. Y.; Wang, Q; Lu, W. Y.; Ju, W; Ahmadian, G; Liu, L; d'Souza, S; Wong, T. P.; Taghibiglou, C; Lu, J; Becker, L. E.; Pei, L; Liu, F; Wymann, M. P.; MacDonald, J. F.; Wang, Y. T. (2003). "Activation of PI3-kinase is required for AMPA receptor insertion during LTP of mEPSCs in cultured hippocampal neurons". Neuron. 38 (4): 611–24. doi:10.1016/s0896-6273(03)00228-9. PMID 12765612.
- ↑ Sui, L; Wang, J; Li, B. M. (2008). "Role of the phosphoinositide 3-kinase-Akt-mammalian target of the rapamycin signaling pathway in long-term potentiation and trace fear conditioning memory in rat medial prefrontal cortex". Learning & Memory. 15 (10): 762–76. doi:10.1101/lm.1067808. PMID 18832563.
- ↑ Sui, L; Wang, J; Li, B. M. (2008). "Role of the phosphoinositide 3-kinase-Akt-mammalian target of the rapamycin signaling pathway in long-term potentiation and trace fear conditioning memory in rat medial prefrontal cortex". Learning & Memory. 15 (10): 762–76. doi:10.1101/lm.1067808. PMID 18832563.