Tandem affinity purification
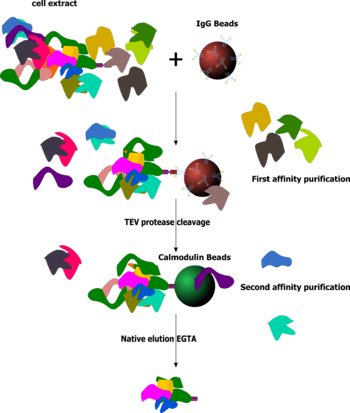
Tandem affinity purification (TAP) is a purification technique for studying protein–protein interactions. It involves creating a fusion protein with a designed piece, the TAP tag, on the end.
In the original version of the technique, the protein of interest with the TAP tag first binds to beads coated with IgG, the TAP tag is then broken apart by an enzyme, and finally a different part of the TAP tag binds reversibly to beads of a different type. After the protein of interest has been washed through two affinity columns, it can be examined for binding partners.
The original TAP method involves the fusion of the TAP tag to the C-terminus of the protein under study. The TAP tag consists of calmodulin binding peptide (CBP) from the N-terminal, followed by tobacco etch virus protease (TEV protease) cleavage site and Protein A, which binds tightly to IgG. The relative order of the modules of the tag is important because Protein A needs to be at the extreme end of the fusion protein so that the entire complex can be retrieved using an IgG matrix.
Many other tag combinations have been proposed since the TAP principle was first published.
Variant tags
This tag is also known as the C-terminal TAP tag because an N-terminal version is also available. However, the method to be described assumes the use of a C-terminal tag, although the principle behind the method is still the same.
History
TAP tagging was invented by a research team working in the European Molecular Biology Laboratory at late 1990s (Rigaut et al., 1999,[1] Puig et al.,2001[2]) and proposed as a new tool for proteome exploration. It was used by the team to characterize several protein complexes (Rigaut et al., 1999,[1] Caspary et al. 1999,[3] Bouveret et al., 2000,[4] Puig et al., 2001[2]). The first large-scale application of this technique was in 2002, in which the research team worked in collaboration with scientists of the proteomics company Cellzome to develop a visual map of the interaction of more than 230 multi-protein complexes in a yeast cell by systematically tagging the TAP tag to each protein. The first successful report of using TAP tag technology in plants came in 2004 (Rohila et al., 2004,[5])
Process
There are a few methods in which the fusion protein can be introduced into the host. If the host is yeast, then one of the methods may be the use of plasmids that will eventually translate the fusion protein within the host. Whichever method that is being used, it is preferable to maintain expression of the fusion protein as close as possible to its natural level.
Once the fusion protein is translated within the host, the new protein at one end of the fusion protein would be able to interact with other proteins. Subsequently, the fusion protein is retrieved from the host by breaking the cells and retrieving the fusion protein through affinity selection, together with the other constituents attached to the new protein, by means of an IgG matrix.
After washing, TEV protease is introduced to elute the bound material at the TEV protease cleavage site. This eluate is then incubated with calmodulin-coated beads in the presence of calcium. This second affinity step is required to remove the TEV protease as well as traces of contaminants remaining after the first affinity step.[2] After washing, the eluate is then released with ethylene glycol tetraacetic acid (EGTA).
The native elution, consisting of the new protein and its interacting protein partners as well as CBP, can now be analyzed by sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) or be identified by mass spectrometry.
Advantages
An advantage of this method is that there can be real determination of protein partners quantitatively in vivo without prior knowledge of complex composition. It is also simple to execute and often provides high yield.[2] One of the obstacles of studying protein protein interaction is the contamination of the target protein especially when we don’t have any prior knowledge of it. TAP offers an effective, and highly specific means to purify target protein. After 2 successive affinity purifications, the chance for contaminants to be retained in the eluate reduces significantly.
Disadvantages
However, there is also the possibility that a tag added to a protein might obscure binding of the new protein to its interacting partners. In addition, the tag may also affect protein expression levels. On the other hand, the tag may also not be sufficiently exposed to the affinity beads, hence skewing the results.
There may also be a possibility of a cleavage of the proteins by the TEV protease, although this is unlikely to be frequent given the high specificity of the TEV protease.[6]
Suitability
As this method involves at least 2 rounds of washing, it may not be suitable for screening transient protein interactions, unlike the yeast two-hybrid method or in vivo crosslinking with photo-reactive amino acid analogs. However, it is a good method for testing stable protein interactions and allows various degrees of investigation by controlling the number of times the protein complex is purified.
Applications
In 2002, the TAP tag was first used with mass spectrometry in a large-scale approach to systematically analyse the proteomics of yeast by characterizing multiprotein complexes.[7] The study revealed 491 complexes, 257 of them wholly new. The rest were familiar from other research, but now virtually all of them were found to have new components. They drew up a map relating all the protein components functionally in a complex network.
Many other proteomic analyses also involve the use of TAP tag. A research by EMBO (Dziembowski, 2004) identified a new complex required for nuclear pre-mRNA retention and splicing. They have purified a novel trimeric complex composed of 3 other subunits (Snu17p, Bud13p and Pml1p) and find that these subunits are not essential for viability but required for efficient splicing (removal of introns) of pre-mRNA. In 2006, Fleischer et al. systematically identified proteins associated with eukaryotic ribosomal complexes.[8] They used multifaceted mass spectrometry proteomic screens to identify yeast ribosomal complexes and then used TAP tagging to functionally link up all these proteins.
Other epitope-tag combinations
The principle of tandem-affinity purification of multiprotein complexes is not limited to the combination of CBP and Protein A tags used in the original work by Rigaut et al. (1999). For example, the combination of FLAG- and HA-tags has been used since 2000 by the group of Nakatani [9][10] to purify numerous protein complexes from mammalian cells. Many other tag combinations have been proposed since the TAP principle was published.
References
- 1 2 Rigaut G, et al. (1999). "A generic protein purification method for protein complex characterization and proteome exploration". Nature Biotechnology. 17 (10): 1030–1032. doi:10.1038/13732. PMID 10504710.
- 1 2 3 4 Puig, O.; et al. (2001). "The Tandem Affinity Purification (TAP) Method: A General Procedure of Protein Complex Purification". Methods. 24 (3): 218–229. doi:10.1006/meth.2001.1183. PMID 11403571.
- ↑ Caspary F, et al. (1999). "Partial purification of the yeast U2 snRNP reveals a novel yeast pre-mRNA splicing factor required for pre-spliceosome assembly". The EMBO Journal. 18 (12): 3463–3474. doi:10.1093/emboj/18.12.3463. PMC 1171425
 . PMID 10369685.
. PMID 10369685. - ↑ Bouveret E, et al. (2000). "A Sm-like protein complex that participates in mRNA degradation". The EMBO Journal. 19 (7): 1661–1671. doi:10.1093/emboj/19.7.1661. PMC 310234. PMID 10747033.
- ↑ Rohila, Jai S.; Chen, Mei; Cerny, Ronald; Fromm, Michael E. (Feb 2004). "Improved tandem affinity purification tag and methods for isolation of protein heterocomplexes from plants". The Plant Journal. 38 (1). doi:10.1111/j.1365-313X.2004.02031.x.
- ↑ Dougherty, W.G., S.M. Cary, and T.D. Parks (1989). "Molecular genetic analysis of a plant virus polyprotein cleavage site: a model". Virology. 171 (2): 356–364. doi:10.1016/0042-6822(89)90603-X. PMID 2669323.
- ↑ Functional organization of the yeast proteome by systematic analysis of protein complexes, Gavin AC et al. Nature 415, 141-147 (10 January 2002) | doi:10.1038/415141a; Received 15 August 2001; Accepted 25 October 2001
- ↑ Systematic identification and functional screens of uncharacterized proteins associated with eukaryotic ribosomal complexes, doi: 10.1101/gad.1422006, Genes & Dev. 2006. 20: 1294-1307
- ↑ Ikura T et al. "Involvement of the TIP60 histone acetylase complex in DNA repair and apoptosis". Cell. 2000 102(4):463-73.
- ↑ Nakatani Y, Ogryzko V. "Immunoaffinity purification of mammalian protein complexes". Methods Enzymol. 2003;370:430-44.