Cell migration
Cell migration is a central process in the development and maintenance of multicellular organisms. Tissue formation during embryonic development, wound healing and immune responses all require the orchestrated movement of cells in particular directions to specific locations. Errors during this process have serious consequences, including intellectual disability, vascular disease, tumor formation and metastasis. An understanding of the mechanism by which cells migrate may lead to the development of novel therapeutic strategies for controlling, for example, invasive tumour cells. Cells often migrate in response to specific external signals, including chemical signals and mechanical signals.[1]
Due to the highly viscous environment (low Reynolds number), cells need to permanently produce forces in order to move. Cells achieve active movement by very different mechanisms. Many less complex prokaryotic organisms (and sperm cells) use flagella or cilia to propel themselves. Eukaryotic cell migration typically is far more complex and can consist of combinations of different migration mechanisms. It generally involves drastic changes in cell shape which are driven by the cytoskeleton. Two very distinct migration scenarios are crawling motion (most commonly studied) and blebbing motility.[2]
Cell migration studies

The migration of cultured cells attached to a surface is commonly studied using microscopy. As cell movement is very slow, a few µm/minute, time-lapse microscopy videos are recorded of the migrating cells to speed up the movement. Such videos (Figure 1) reveal that the leading cell front is very active with a characteristic behavior of successive contractions and expansions. It is generally accepted that the leading front is the main motor that pulls the cell forward.
Common features
The processes underlying mammalian cell migration are believed to be consistent with those of (non-spermatozooic) locomotion.[4] Observations in common include:
- cytoplasmic displacement at leading edge (front)
- laminar removal of dorsally-accumulated debris toward trailing edge (back)
The latter feature is most easily observed when aggregates of a surface molecule are cross-linked with a fluorescent antibody or when small beads become artificially bound to the front of the cell.[5]
Other eukaryotic cells are observed to migrate similarly. The amoeba Dictyostelium discoideum is useful to researchers because they consistently exhibit chemotaxis in response to cyclic AMP; they move more quickly than cultured mammalian cells; and they have a haploid genome that simplifies the process of connecting a particular gene product with its effect on cellular behaviour.


Molecular processes of migration
There are two main theories for how the cell advances its front edge: the cytoskeletal model and membrane flow model. It is possible that both underlying processes contribute to cell extension.
Leading Edge
Experimentation has shown that there is rapid actin polymerisation at the cell's front edge.[6] This observation has led to the hypothesis that formation of actin filaments "push" the leading edge forward and is the main motile force for advancing the cell’s front edge.[7][8] In addition, cytoskeletal elements are able to interact extensively and intimately with a cell's plasma membrane.[9]
Trailing edge
Other cytoskeletal components (like microtubules) have important functions in cell migration. It has been found that microtubules act as “struts” that counteract the contractile forces that are needed for trailing edge retraction during cell movement. When microtubules in the trailing edge of cell are dynamic, they are able to remodel to allow retraction. When dynamics are suppressed, microtubules cannot remodel and, therefore, oppose the contractile forces.[10] The morphology of cells with suppressed microtubule dynamics indicate that cells can extended the front edge (polarized in the direction of movement), but have difficulty retracting their trailing edge.[11] On the other hand high drug concentrations, or microtubule mutations that depolymerize the microtubules, can restore cell migration but there is a loss of directionality. It can be concluded that microtubules act both to restrain cell movement and to establish directionality (see "Polarity in Migrating Cells" below).
Membrane flow model (B)
Studies have also shown that the front is the site at which membrane is returned to the cell surface from internal membrane pools at the end of the endocytic cycle.[12] This has led to the hypothesis that extension of the leading edge occurs primarily by addition of membrane at the front of the cell. If so, the actin filaments that form at the front might stabilize the added membrane so that a structured extension, or lamella, is formed rather than a bubble-like structure (or bleb) at its front.[13] For a cell to move, it is necessary to bring a fresh supply of "feet" (proteins called integrins, which attach a cell to the surface on which it is crawling) to the front. It is likely that these feet are endocytosed toward the rear of the cell and brought to the cell's front by exocytosis, to be reused to form new attachments to the substrate.
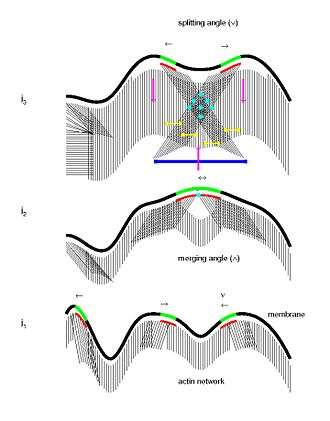
Collective biomechanical and molecular mechanism of cell motion
Based on some mathematical models, recent studies hypothesize a novel biological model for collective biomechanical and molecular mechanism of cell motion.[14] It is proposed that microdomains weave the texture of cytoskeleton and their interactions mark the location for formation of new adhesion sites. According to this model, microdomain signaling dynamics organizes cytoskeleton and its interaction with substratum. As microdomains trigger and maintain active polymerization of actin filaments, their propagation and zigzagging motion on the membrane generate a highly interlinked network of curved or linear filaments oriented at a wide spectrum of angles to the cell boundary. It is also proposed that microdomain interaction marks the formation of new focal adhesion sites at the cell periphery. Myosin interaction with the actin network then generate membrane retraction/ruffling, retrograde flow, and contractile forces for forward motion. Finally, continuous application of stress on the old focal adhesion sites could result in the calcium-induced calpain activation, and consequently the detachment of focal adhesions which completes the cycle.
Polarity in migrating cells
Migrating cells have a polarity—a front and a back. Without it, they would move in all directions at once, i.e. spread. How this polarity is formulated at a molecular level inside a cell is unknown. In a cell that is meandering in a random way, the front can easily give way to become passive as some other region, or regions, of the cell form(s) a new front. In chemotaxing cells, the stability of the front appears enhanced as the cell advances toward a higher concentration of the stimulating chemical. This polarity is reflected at a molecular level by a restriction of certain molecules to particular regions of the inner cell surface. Thus, the phospholipid PIP3 and activated Rac and CDC42 are found at the front of the cell, whereas Rho GTPase and PTEN are found toward the rear.[15][16]
It is believed that filamentous actins and microtubules are important for establishing and maintaining a cell’s polarity. Drugs that destroy actin filaments have multiple and complex effects, reflecting the wide role that these filaments play in many cell processes. It may be that, as part of the locomotory process, membrane vesicles are transported along these filaments to the cell’s front. In chemotaxing cells, the increased persistence of migration toward the target may result from an increased stability of the arrangement of the filamentous structures inside the cell and determine its polarity. In turn, these filamentous structures may be arranged inside the cell according to how molecules like PIP3 and PTEN are arranged on the inner cell membrane. And where these are located appears in turn to be determined by the chemoattractant signals as these impinge on specific receptors on the cell’s outer surface.
Although microtubules have been known to influence cell migration for many years, the mechanism by which they do so has remained controversial. On a planar surface, microtubules are not needed for the movement, but they are required to provide directionality to cell movement and efficient protrusion of the leading edge.[11][17] When present, microtubules retard cell movement when their dynamics are suppressed by drug treatment or by tubulin mutations.[11]
Inverse problems in the context of cell motility
In a series of recent works a new area of research called inverse problems in cell motility has been established. [18] [19][14] This approach is based on the idea that behavioral or shape changes of a cell bear information about the underlying mechanisms that generate these changes. Reading cell motion, namely, understanding the underlying biophysical and mechanochemical processes, is of paramount importance. [20] [21] The mathematical models developed in these works determine some physical features and material properties of the cells locally through analysis of live cell image sequences and uses this information to make further inferences about the molecular structures, dynamics, and processes within the cells, such as the actin network, microdomains, chemotaxis, adhesion, and retrograde flow.
See also
- Cap formation
- Chemotaxis
- Durotaxis
- Endocytic cycle
- Neurophilia
- Mouse models of breast cancer metastasis
External links
- Cell Migration Gateway The Cell Migration Gateway is a comprehensive and regularly updated resource on cell migration
- The Cytoskeleton and Cell Migration A tour of images and videos by the J. V. Small lab in Salzburg and Vienna
- Time-lapse microscopy videos showing proliferating and migrating cells
- Cell migration tracking by Phase Holographic Imaging AB
References
- ↑ Mak, M.; Spill, F.; Roger, K.; Zaman, M. "Single-Cell Migration in Complex Microenvironments: Mechanics and Signaling Dynamics". Journal of biomechanical engineering. 138: 021004. doi:10.1115/1.4032188.
- ↑ Huber, F; Schnauss, J; Roenicke, S; Rauch, P; Mueller, K; Fuetterer, C; Kaes, J (2013). "Emergent complexity of the cytoskeleton: from single filaments to tissue". Advances in Physics. 62 (1): 1–112. doi:10.1080/00018732.2013.771509. PMID 24748680. online
- ↑ "HoloMonitor - Non-invasive image cytometers". Phase Holographic Imaging AB.
- ↑ "What is Cell Migration?". Cell Migration Gateway. Cell MIgration Consortium. Retrieved 24 March 2013.
- ↑ Abercrombie, M; Heaysman, JE; Pegrum, SM (1970). "The locomotion of fibroblasts in culture III. Movements of particles on the dorsal surface of the leading lamella". Experimental Cell Research. 62 (2): 389–98. doi:10.1016/0014-4827(70)90570-7. PMID 5531377.
- ↑ Wang, Y. L. (1985). "Exchange of actin subunits at the leading edge of living fibroblasts: possible role of treadmilling". The Journal of Cell Biology. 101 (2): 597–602. doi:10.1083/jcb.101.2.597. PMC 2113673
 . PMID 4040521.
. PMID 4040521. - ↑ Mitchison, T; Cramer, LP (1996). "Actin-Based Cell Motility and Cell Locomotion". Cell. 84 (3): 371–9. doi:10.1016/S0092-8674(00)81281-7. PMID 8608590.
- ↑ Pollard, Thomas D; Borisy, Gary G (2003). "Cellular Motility Driven by Assembly and Disassembly of Actin Filaments". Cell. 112 (4): 453–65. doi:10.1016/S0092-8674(03)00120-X. PMID 12600310.
- ↑ Doherty, Gary J.; McMahon, Harvey T. (2008). "Mediation, Modulation, and Consequences of Membrane-Cytoskeleton Interactions". Annual Review of Biophysics. 37: 65–95. doi:10.1146/annurev.biophys.37.032807.125912. PMID 18573073.
- ↑ Yang, Hailing; Ganguly, Anutosh; Cabral, Fernando (2010). "Inhibition of Cell Migration and Cell Division Correlates with Distinct Effects of Microtubule Inhibiting Drugs". The Journal of Biological Chemistry. 285 (42): 32242–50. doi:10.1074/jbc.M110.160820. PMC 2952225. PMID 20696757.
- 1 2 3 Ganguly, A; Yang, H; Sharma, R; Patel, K; Cabral, F (2012). "The Role of Microtubules and Their Dynamics in Cell Migration.". J Biol Chem. 4 (52): 253–65. doi:10.1074/jbc.M112.423905. PMID 23135278.
- ↑ Bretscher, M. S. (1983). "Distribution of receptors for transferrin and low density lipoprotein on the surface of giant HeLa cells". Proceedings of the National Academy of Sciences. 80 (2): 454–8. doi:10.1073/pnas.80.2.454. PMC 393396. PMID 6300844.
- ↑ Bretscher, M (1996). "Getting Membrane Flow and the Cytoskeleton to Cooperate in Moving Cells". Cell. 87 (4): 601–6. doi:10.1016/S0092-8674(00)81380-X. PMID 8929529.
- 1 2 3 Coskun, Hasan; Coskun, Huseyin. (March 2011). "Cell physician: reading cell motion. A mathematical diagnostic technique through analysis of single cell motion". Bull Math Biol. 73 (3): 658–82. doi:10.1007/s11538-010-9580-x.
- ↑ Parent, C. A.; Devreotes, PN (1999). "A Cell's Sense of Direction". Science. 284 (5415): 765–70. doi:10.1126/science.284.5415.765. PMID 10221901.
- ↑ Ridley, A. J.; Schwartz, MA; Burridge, K; Firtel, RA; Ginsberg, MH; Borisy, G; Parsons, JT; Horwitz, AR (2003). "Cell Migration: Integrating Signals from Front to Back". Science. 302 (5651): 1704–9. doi:10.1126/science.1092053. PMID 14657486.
- ↑ Meyer, A.S.; Hughes-Alford, S.K.; Kay, J.E.; Castillo, A.; Wells, A.; Gertler, F.B.; Lauffenburger, D.A. (2012). "2D protrusion but not motility predicts growth factor–induced cancer cell migration in 3D collagen". J Cell Biol. 194 (6): 721–729. doi:10.1083/jcb.201201003. PMC 3373410. PMID 22665521.
- ↑ Coskun, Huseyin. (2006). Mathematical Models for Ameboid Cell Motility and Model Based Inverse Problems. ProQuest.
- ↑ Coskun, Huseyin; Li, Yi; Mackey, Mackey A. (Jan 2007). "Ameboid cell motility: a model and inverse problem, with an application to live cell imaging data". J Theor Boil. 244 (2): 169–79. doi:10.1016/j.jtbi.2006.07.025.
- ↑ "Profiling Cells with Math". Mathematical Association of America.
- ↑ "Mathematicians use cell 'profiling' to detect abnormalities -- including cancer". ScienceDaily.
