Resolvin
Resolvins are autacoids of a specific lipid structure: dihydroxy or trihydroxy metabolites of omega-3 fatty acids, primarily eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA) but also the docosapentaenoic acid (DPA), clupanodonic acid. They are members of an expanding class of polyunsaturated fatty acid (PUFA) metabolites termed specialized proresolving mediators (SPMs). Other SPMs include the lipoxins, protectin D1 and its related products, and the maresins. SPMs are locally formed and locally acting cell signaling autocoids: they are made by cells and act upon their parent or nearby cells to coordinate functional responses. SPMs possess potent anti-inflammation, tissue protection, and tissue healing activities in diverse animal models and accordingly are proposed to be involved in resolving physiological inflammatory responses. Their failure to form in adequate amounts is also proposed to underlie a broad range of human diseases involving pathological inflammation.[1][2][3][4] Metabolically stable analogs of the SPMs, including the resolvins, are in development and being tested in volunteers with chronic inflammation-related diseases.
Biochemistry and production
Resolvins (Rvs) fall into several sub-classes based on the straight chain PUFA from which they are formed and/or a unique aspect of their structure. The Resolvin Ds (RvDs) are metabolites of the 22-carbon PUFA, DHA (i.e. 4Z,7Z,10Z,13Z,16Z,19Z)-docosahexaenoic acid); the resolvin Es (RvEs) are metabolites of the 20-carbon PUFA, EPA (i.e. 5Z,8Z,11Z,14Z,17Z-5,8,11,14,17-eicosapentaenoic acid); the resolvin Dn-6DPA (RvDsn-6DPA) are metabolites of the DPA isomer, osbond acid (i.e. 4Z,7Z,10Z,13Z,16Z-docosapentaenoic acid; the resolvin Dn-3DPA (RvDn-3DPA) are metabolites of the DPA isomer, clupanodonic acid (i.e. 7Z,10Z,13Z,16Z,19Z)-docosapentaenoic acid); and the resolvin Ts (RvTs) are metabolites of clupanodonic acid that, in contrast to (RvDsn-3DPA (all of which possess a 17S-hydroxyl residue), possess a 17R-hydroxyl residue. Certain isomers of RvDs are termed aspirin-triggered resolvin Ds (AT-RvDs) because their synthesis is initiated by a drug-modified COX2 enzyme to form 17(R)-hydroxyl rather than 17(S)-hydroxyl residue of the ReVEs; however, an as yet unidentified cytochrome P450 enzyme(s) may also forms this 17(R)-hydroxy intermediate and thereby contribute to the production of AT-RvEs. All of the cited resolvins except the RvDsn-6DPAs are metabolites of omega-3 fatty acids; it is proposed that the metabolism of dietary omega-3 fatty acids to these resolvins is an important mechanism by which these diets may ameliorate diverse inflammatory reactions and inflammation-based diseases.[1][2][3][4]
The following oxygenase enzymes are responsible for metabolizing PUFA to resolvins: 15-lipoxygenase-1 (i.e. ALOX15), possibly 15-lipoxygenase-2 (i.e. ALOX15B), 5-lipoxygenase (i.e. ALOX5), cyclooxygenase-2 (i.e. COX-2), and certain Cytochrome P450 monooxygenases.[3][4] While only one of these enzymes may form certain Rvs, more commonly two different oxygenases act in series to do so; in the latter cases, each oxygenase often resides in a different cell type with one cell passing along the intermediate metabolite that it forms to the other cell type. As an example of this "transcellular metabolic pathway", aspirin triggered resolvin Ds (AT-RvDs) re made by the initial conversion of DHA by aspirin-acetylated COX-2 in vascular endothelial cells to 17(R)-hydroxy-DHA which is then transferred to nearby neutrophils which then use ALOX5 to oxygenate the 17(R)-hydroxyl-DHA to AT-resolvin Ds.[4][5]
Resolvin Ds
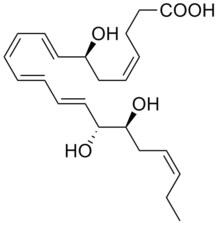
RvDs are poly-hydroxyl metabolites of DHA. To date, six RvD's, which vary in the number, position, and chirality of their hydroxyl residues as well as the position and cis-trans isomerism of their 6 double bonds, have been described. These are: RvD1 (7S,8R,17S-trihydroxy-DHA), RvD2 (7S,16R,17S-trihydroxy-DHA), RvD3 (4S,7R,17S-trihydroxy-DHA), RvD4 (4S,5,17S-trihydroxy-DHA; chirality at position 5 not yet determined), RvD5 (7S,17S-dihydroxy-DHA), and RvD6 (4S,17S-dihydroxy-DHA). (The structures of these RvDs are further defined at Specialized proresolving mediators#DHA-derived Resolvins). These metabolites are formed by a wide range of cells and tissues by the initial metabolism of DHA to 7S-hydroperoxy-DHA and 4S-hydroperoxy-DHA by a 15-lipoxygenase (either ALOX15 or possibly ALOX15B) followed by the further metabolism of the two intermediates by ALOX5 to their 17-hydroperoxy derivatives; these di-hydroperoxy products are further altered to the cited RvDs by these oxygenases or by non-enzymatic reactions and the conversion of their peroxy residues ubiquitous cellular peroxidases.[5][6]
14S,20R-dihyrdoxy-4Z,7Z,10Z,12E,16Z,18E-docosahexaenoic acid, while not yet assigned a RvD number, qualifies as a RvD. It is a DHA metabolite made by mouse eosinophils, detected in the peritoneal fluid of mice undergoing experimental peritonitis, and possessing the ability to inhibit the influx of leukocytes into the peritoneum of mice undergoing experimental peritonitis.[7][8]
AT-resolvins Ds
Aspirin-Triggered resolving D's (AT-RvD's) have structures identical to those of the RvDs except that they have a 17R rather than 17S hydroxyl residue: RvD1 (7S,8R,17R-trihydroxy-DHA), RvD2 (7S,16R,17R-trihydroxy-DHA), RvD3 (4S,7R,17R-trihydroxy-DHA), RvD4 (4S,5,17R-trihydroxy-DHA; chirality at position 5 not yet determined), RvD5 (7S,17R-dihydroxy-DHA), and RvD6 (4S,17R-dihydroxy-DHA). (The structures of these RvDs are further defined at Specialized proresolving mediators#DHA-derived Resolvins.) AT-RvD's are formed under special circumstances; COX-2 modified in activity by aspirin or atorvastatin (a statin-type drug) or, alternatively, a microbial or possibly mammalian cytochrome P450 monoxygenase metabolizes DHA to its 17R-hydroperoxy derivative; this intermediate is then further metabolized as described for the 17S-hydroperoxy intermediate in the previous paragraph to form the cited AT-RvD products.[3] RvD1 (7S,8R,17S-trihydroxy-DHA), RvD2 (7S,16R,17S-trihydroxy-DHA), RvD3 (4S,7R,17S-trihydroxy-DHA), RvD4 (4S,5,17S-trihydroxy-DHA; chirality at position 5 not yet determined), RvD5 (7S,17S-dihydroxy-DHA), and RvD6 (4S,17S-dihydroxy-DHA). (The structures of these RvDs are further defined at Specialized proresolving mediators#DHA-derived Resolvins).
Resolvin Es
RvEs are di- or tri-hydroxyl metabolites of EPA. To date, four RvE's have been described: RvE1 (5S,12R,18R-trihydroxy-EPA), 18S-Rv1 (5S,12R,18S-trihydroxy-EPA), RvE2 (5S,18R-dihydroxy-EPA), and RvE3 (17R,18R/S-dihydroxy-EPA). (Structures of the RvEs are further defined at Specialized proresolving mediators#EPA-derived Resolvins.) The chirality of the hydroxyl residue at carbon 18 for RvE3 has not been determined; clearly one but possibly both of the 18R- and 18S R-RvD3 diastereoisomers is/are active. Resolvin Es are formed in manner similar to AT resolving T's. COX-2 modified in activity by aspirin or atorvastatin or, alternatively, a microbial or possibly mammalian cytochrome P450 monoxygenase metabolizes EPA to its 18R-hydroperoxy derivative; this intermediate is then further metabolized by ALOX5 to a 5,6 epoxide which is hydrolyzed enzymatically or non-enzymatically to RvE1 and 18S-RvE1 or reduced to RvE2; alternatively the 18R-hydroperoxide is converted to the 17R,18S vicinal diol product, RvE3.[5][6]
T series resolvins
T series resolvins are very recently described resolvins. In the model system used to identify them, human platelets pretreated with aspirin or atorvastatin metabolize the omega-3 DPA, clupanodonic acid (DPAn-3 ) by aspirin-treated or atorvastatin-treated COX2 to a 13S-hydroperoxy intermediate (aspirin and atorvastatin change the activity of COX2 from a cyclooxygenase to an hydroxyperoxidase-forming enzyme. The intermediate is then passed to nearby human neutrophils which metabolize it, probably by ALOX5 enzyme activity, to four poly-hydroxyl metabolites: RvT1 (7,13R,20-trihydroxy-DPAn-3); RvT2 (7,8,13R-trihydroxy-DPAn-3); RvT3 (7,12,13R-trihydroxy-8Z,10E,14E,16Z,19Z-DPAn-3) and RvT4 (7,13R-dihydroxy-DPAn-3). The chirality of the 13 hydroxyl residue is assumed but not yet formally proven; the chirality of the other hydroxyl residues has not yet been determined. Subsequent studies found that these four RvT's are also formed by mixtures of human neutrophils and vascular endothelium cells and, additionally, are detected in the infected tissues of rodents and humans.[9][10][7]
Resolvin Dn-3DPA
RvDn-3DPA are di- and tri-hydroxyl metabolites of DPAn-3DPA, i.e. clupanodonic acid. Three RvDn-3DPA have been described: RvD1n-3DPA (7,8,17-trihydroxy-DPAn-3DPA), RvD2n-3DPA (7,16,17-trihydroxy-DPAn-3DPA), and RvD5n-3DPA. The chirality of these hydroxyl residues has not yet been defined (other structural details of the RvDn-3DPA are given at Specialized proresolving mediators#n-3 DPA-derived resolvins). Cells form these resolvins by metabolizing clupanodonic acid, presumptively with a 15-lipoxygenase enzyme, to form 17S-hydroperoxy intermediate which is further converted to a 7S,17S-dihydroperoxy intermediate possibly by ALOX5. The latter intermediate may be reduced by cellular peroxidases to RvD5n-3DPA or, alternatively, convert to a 7,8-epoxide or 16,17-epoxide which, after reduction of their hydroperoxide to hydroxyl residues, then form the vicinal diol tri-hydroxyl products, RvD1n-3DPA and RvD2n-3DPA, respectively.[2]
Mechanisms of Action
The mechanism(s) by which each of the resolvins activate cells has not been fully elucidated. However, many resolvins appear to operate at least in part by acting through the following G protein-coupled receptors (GPRs): 1) RvD1 and AT-RvD1 act through the Formyl peptide receptor 2, which is also activated by certain lipoxins and is therefore often termed the ALX/FPR2 receptor; 2) RvD1, AT-RVD1, RvD3, AT-RvD3, and RvD5 act through the GPR32 receptor which is now also termed the RVD1 receptor; 3) RvD2 acts through the GPR18 receptor also now termed the RvD2 receptor; GPR18 is also a receptor for N-Arachidonylglycine, Anandamide, Arachidonylcyclopropylamide, Abnormal cannabidiol, Δ9-Tetrahydrocannabinol, and other agents (see GPR18; and 4) RvE1 and the 18(S) analog of RvE1 are full activators while RvE2 is a partaial actiator of the CMKLR1 receptor which is also known as the chemR23 or Chemokine-like receptor 1; CMKLR1 is also activated by chemerin and Adipokines. All of these receptors activate their parent cells through standard GPR-mobilized pathways (see G protein–coupled receptor# G-protein-dependent signaling) to elicit their anti-inflammatory functions.[4][11] RvE1, 18(S)-RvE1, and RvE2 inhibit the Leukotriene B4 receptor 1 which is the receptor for inflammation-promoting PUFA metabolites such as LTB4 and the R stereoisomer of 12-HETE; by inhibiting the action of these pro-inflammatory mediators, these SRM's contribute to inhibiting inflammation.[5][11][4]
The resolvins also effect receptors that are not GPRs: a) RvD1 also inhibits activation of TRPV3, TRPV4, and TRPA1 receptors; b) RvD2 inhibits activation of TRPV1 and TRPA1 receptors; c) AT-RvD1 inhibits activation of the TRPV3 receptor; and d) ReV1 inhibits activation of the TRPV1 and TRPV3 receptors. These receptors are located on nerve terminals of neurons in the peripheral nervous system, on dorsal root ganglia, and/or on spinal cord; they are involved in the perception of mechanical pain (TRPA1 and TRPV4), heat pain (TRPV3 and TRPV4), and hypotonicity-related pain. The mechanism behind the resolvin's inhibitory effects on them in not clear but may be indirect and involve at least in certain cases the activation of a GPRs located on the neruons or nearbymicroglia and/or astrocyte cells of the nerve tissue. CMKLR1, GPR32, and FPR2 have been proposed to be the GPR through which these resolvins act.[12] RvE1 also inhibits activation of the NMDA receptor, a nerve cell non-GPR receptor and ion channel which is activated by glutamate, glycine, and D-serine; this receptor is thought to be critical in synaptic plasticity, a cellular mechanism for learning and memory that also may be involved in the development of chronic pain. The mechanisms by which the non-GPR receptors act can be found on the cited Wikipedia pages.[5][6][12][13][14]
Activities and functions
A prevailing theory holds that inflammation provoking insults lead to the production of arachidonic acid metabolites (e.g. prostaglandins, leukotrienes, and 5-oxo-eicosatetraenoic acids) and various cytokines (e.g. Interleukin 8, Interleukin 2, and Granulocyte macrophage colony-stimulating factor) that orchestrate the ensuing innate immunity-based inflammatory responses. Later in these responses, production of the cited types of arachidonic acid metabolites switches to the production of SPMs, i.e. the resolvin metabolites of EPA, DHA, and DPA and the Maresin and Protectin D1 metabolites of DHA. The SPMs proceed to resolve these responses and initiate healing.[11]
The most thoroughly studied resolvins, RvE1 and RvD1: a) inhibit blood leukocytes (e.g. neutrophils and eosinophils) from migrating out of the circulation into sites of inflammation and allergy; b) stimulate macrophages to convert from a M1-like pro-inflammatory phenotype to a tissue repairing and wound healing M2 phenotype (see macrophage#Macrophage subtypes; c) stimulate macrophage phagocytosis ((i.e. ingestion) of apoptotic (i.e. dying) leukocytes at tissue sites of inflammation and allergy in a physiological process termed Efferocytosis; d) stimulate Natural killer T cells to clear leukocytes from inflamed tissues; e) stimulate leukocytes to engulf (i.e. eat) and kill pathogenic microbes at sites of invasion; f) inhibit the production of pro-inflammatory cytokines (e.g. TNFα, Interleukin 1β, Interleukin 6, Interleukin 8, Interleukin 12, et.) and chemokines (e.g. CXCL2, CXCL8, CCL5, etc.) while stimulating the production of anti-inflammatory cytokines (e.g. Interleukin 10); g) inhibit activation and function of dendritic cells, activation (e.g. aggregation) of platelets, and migration of vascular smooth muscle; and h) promote tissue repair and healing.[4] In consequence of these activities, RvE1 and RvD1 prevent or reduce the severity, tissue destruction, or mortality in animal models of a diverse set inflammatory and allergic reactions to bacterial pathogens, mechanical injuries, burns, cigarette smoke, autoimmune reactions, and various neurological and non-neurological degenerative diseases.[4][15] These animal model studies suggest that RvE1 and RvD1 contribute to resolving not only standard inflammatory and allergic reactions and diseases but also may be involved in slowing or, if deficient, promoting the progression of Alzheimer's disease, fibromyalgia, exercise-induced muscle injury, atherosclerosis, neuralgias, obesity, Retinopathy involving neovascularization, and, by its presence in mothers' milk, protection of suckling infants.[4][16][17][18][19] Preclinical studies finding that human cells and issues in culture respond to and show the presence and increases in the levels of the resolvins as well as findings that RvE1 RvD1 are found at increased or in other instances decreased levels in the fluids and/or tissues of humans suffering the cited diseases lend further support to the notion that their presence slows while their absence promotes such diseases in humans.[2][3][20][21] RvD1 also inhibits activation of the TRPV3, TRPV4, and TRPA1 receptors and ReV1 inhibits activation of the TRPV1 and TRPV3 receptors. this inhibition of these nervous tissue receptors appears responsible for reducing pain perception in various animal models and suggests that the two resolves may also function to reduce pain in humans.[5][13][12]
In a mouse model of sepsis caused by caecal ligation and puncture, RvD2 decreased local and systemic bacterial burden, excessive cytokine production and neutrophil recruitment, while increasing peritoneal mononuclear cells and macrophage phagocytosis as well as animal survival; in a mouse model of burn-related sepsis, RvD2 likewise increased survival when mice were injected with a lethal amount of endotoxin.[22] RvD2 also reduced neutrophil accumulation, plasma levels of tumor necrosis factor-α, and plasma levels of granulocyte macrophage colony-stimulating factor while increasing perfusion recovery as well as the number of regenerating myocytes in injured muscle tissue in a mouse model of hind limb ischemia.[23]
RvD5 reduced blood levels of bacteria, serum levels of the pro-inflammatory cytokines TNF-α and IL-1β, and a symptom of shock in a mouse models of E. coli-induced peritonitis. In Staphylococcus aureus-induced skin infection using mouse dorsal skin pouches, RvD5 enhanced the clearance of bacteria while lowering viable bacterial titers and reducing neutrophil infiltration in pouch exudates. These effects were enhanced when either RvD1 or the antibiotic, vancomycin, was administered along with Rvd5 to the mice.[22]
While all of the resolvins possess one or more anti-inflammatory activity as accessed on in vitro model systems, not all of the resolvins have been studied for activity in vivo as long or as thoroughly as RvE1 and RvD1. RvD2 and to lesser extents AT-RvD1 and RvD5 have been found effective in blocking inflammation in several animal models while RvE2, RvE3, 18S-RvE3, and RvD3 are effective in blocking inflammation in one or two animal models.[12][5][6][19] The recently defined SPMs, the RvTs, have potent in vitro and in vivo anti-inflammatory activity, being particularly effective in reducing the systemic inflammation as well as increasing the survival in a mouse animal model injected with lethal doses of E. coli bacteria.[10][7]
Resolvins as mediators of omega-3 fatty acids
Several studies have suggested that the anti-inflammatory and other beneficial effects of omega-3 fatty acid-rich diets are due at least in part to their metabolic conversion to resolvins. However, a recent translational study questions the notion that resolvins and other members of the SPM family are indeed formed in the human body from omega-3 PUFA. This study failed to detect a consistent signal of resolvin formation in urine or plasma of healthy volunteers who had taken fish oil. This study also found no alteration in the formation of resolvins during the resolution of inflammation which was induced by bacterial lipopolysaccharide in these volunteers. By contrast, formation of a series of established enzymatic and nonenzymatic oxidation products formed from omega-3 PUFA could readily be demonstrated in vivo. On this basis, the study authors concluded that their study fails to provide evidence consistent with the hypothesis that resolvins mediate an anti-inflammatory action of fish oil.[24] Further information can be found in a commentary accompanying this translational work.[25] This commentary indicated that the area of dietary omega-3 PUFA in the prevention and amelioration of inflammatory diseases remains somewhat clouded with contradictory results, controversial, and in need of further rigorous study. It will be important to establish the production and presence of resolvins and other SPMs given their potent protective actions in humans. In this regard, using rigorous and validated mass spectrometry based methods many human tissues and cell types have been shown to produce SPM in ror example, human white blood cells such as macrophages,[26] serum, lymph nodes, and spleen,[27] urine,[28] placenta,[29] diseased kidney,[30] and, importantly, in milk from lactating mothers.[17] Expert investigators in this field of resolution biology and physiology are now able to simultaneoulsy identify and measure both pro-inflammatory and anti-inflammatory-pro-resolving mediators such as the SPM in human tissues. Thus in the near future it is very likely that lipid mediator and SPM signature profiles will be useful in both precision and personalized medicine.
Clinical studies
Regardless of their presence and function relative to omega-3 PUFA as well as to human disease, the resolvins and other SPMs, or more particularly, their pharmacological analogs that resist being metabolized and therefore exhibit longer life and greater potency in vivo may find clinical utility in suppressing a wide range of human diseases and untoward reactions. To date, however, only one clinical development study on a resolvin or resolvin analog has been reported: the RvE1 analog, RX-10045, significantly improved signs and symptoms in a phase 2 clinical trial of patients with Keratoconjunctivitis sicca (i.e. dry eye syndrome).[3] The follow-up phase III clinical trial is in progress (Safety and Efficacy Study of RX-10045 on the Signs and Symptoms of Dry Eye, identifier NCT00799552; www.clinical trials.gov). A single clinical study on another type of SPM is relevant here: the 15(R/S)-methyl-LXA4 analog of LXA4 significantly reduced the severity of eczema in a double-blind, placebo-controlled, randomized, parallel-groups comparative study of 60 patients.[3][31] Many more clinical studies will be needed to define the efficacy of such analogs in human disease.
External links
- 1 2 Shinohara, M; Mirakaj, V; Serhan, C. N. (2012). "Functional Metabolomics Reveals Novel Active Products in the DHA Metabolome". Frontiers in Immunology. 3: 81. doi:10.3389/fimmu.2012.00081. PMC 3342038
 . PMID 22566962.
. PMID 22566962. - 1 2 3 4 Weylandt KH (2015). "Docosapentaenoic acid derived metabolites and mediators - The new world of lipid mediator medicine in a nutshell". European Journal of Pharmacology. doi:10.1016/j.ejphar.2015.11.002. PMID 26546723.
- 1 2 3 4 5 6 7 Serhan, C. N.; Chiang, N; Dalli, J; Levy, B. D. (2014). "Lipid mediators in the resolution of inflammation". Cold Spring Harbor Perspectives in Biology. 7 (2): a016311. doi:10.1101/cshperspect.a016311. PMID 25359497.
- 1 2 3 4 5 6 7 8 9 Duvall, M. G.; Levy, B. D. (2015). "DHA- and EPA-derived resolvins, protectins, and maresins in airway inflammation". European Journal of Pharmacology. doi:10.1016/j.ejphar.2015.11.001. PMC 4854800. PMID 26546247.
- 1 2 3 4 5 6 7 Qu Q, Xuan W, Fan GH (2015). "Roles of resolvins in the resolution of acute inflammation". Cell Biology International. 39 (1): 3–22. doi:10.1002/cbin.10345. PMID 25052386.
- 1 2 3 4 Serhan CN, Chiang N, Dalli J, Levy BD (2015). "Lipid mediators in the resolution of inflammation". Cold Spring Harbor Perspectives in Biology. 7 (2): a016311. doi:10.1101/cshperspect.a016311. PMID 25359497.
- 1 2 3 Barden AE, Mas E, Mori TA (2016). "n-3 Fatty acid supplementation and proresolving mediators of inflammation". Current Opinion in Lipidology. 27 (1): 26–32. doi:10.1097/MOL.0000000000000262. PMID 26655290.
- ↑ Yokokura Y, Isobe Y, Matsueda S, Iwamoto R, Goto T, Yoshioka T, Urabe D, Inoue M, Arai H, Arita M (2014). "Identification of 14,20-dihydroxy-docosahexaenoic acid as a novel anti-inflammatory metabolite". Journal of Biochemistry. 156 (6): 315–21. doi:10.1093/jb/mvu044. PMID 25012818.
- ↑ Dalli J, Colas RA, Serhan CN (2013). "Novel n-3 immunoresolvents: structures and actions". Scientific Reports. 3: 1940. doi:10.1038/srep01940. PMC 3672887. PMID 23736886.
- 1 2 Dalli J, Chiang N, Serhan CN (2015). "Elucidation of novel 13-series resolvins that increase with atorvastatin and clear infections". Nature Medicine. 21 (9): 1071–5. doi:10.1038/nm.3911. PMC 4560998. PMID 26236990.
- 1 2 3 Serhan, C. N. (2014). "Pro-resolving lipid mediators are leads for resolution physiology". Nature. 510 (7503): 92–101. doi:10.1038/nature13479. PMC 4263681. PMID 24899309.
- 1 2 3 4 Lim JY, Park CK, Hwang SW (2015). "Biological Roles of Resolvins and Related Substances in the Resolution of Pain". BioMed Research International. 2015: 830930. doi:10.1155/2015/830930. PMC 4538417. PMID 26339646.
- 1 2 Ji RR, Xu ZZ, Strichartz G, Serhan CN (2011). "Emerging roles of resolvins in the resolution of inflammation and pain". Trends in Neurosciences. 34 (11): 599–609. doi:10.1016/j.tins.2011.08.005. PMC 3200462. PMID 21963090.
- ↑ Serhan CN, Chiang N, Dalli J (2015). "The resolution code of acute inflammation: Novel pro-resolving lipid mediators in resolution". Seminars in Immunology. 27 (3): 200–15. doi:10.1016/j.smim.2015.03.004. PMC 4515371. PMID 25857211.
- ↑ Schwanke, R. C.; Marcon, R; Bento, A. F.; Calixto, J. B. (2015). "EPA- and DHA-derived resolvins' actions in inflammatory bowel disease". European Journal of Pharmacology. doi:10.1016/j.ejphar.2015.08.050. PMID 26325092.
- ↑ Hersberger, M (2010). "Potential role of the lipoxygenase derived lipid mediators in atherosclerosis: Leukotrienes, lipoxins and resolvins". Clinical Chemistry and Laboratory Medicine. 48 (8): 1063–73. doi:10.1515/CCLM.2010.212. PMID 20441482.
- 1 2 Weiss, Gisela Adrienne; Troxler, Heinz; Klinke, Glynis; Rogler, Daniela; Braegger, Christian; Hersberger, Martin (2013-06-15). "High levels of anti-inflammatory and pro-resolving lipid mediators lipoxins and resolvins and declining docosahexaenoic acid levels in human milk during the first month of lactation". Lipids in Health and Disease. 12 (1): 89. doi:10.1186/1476-511X-12-89. ISSN 1476-511X. PMC 3698171. PMID 23767972.
- ↑ Lim, J. Y.; Park, C. K.; Hwang, S. W. (2015). "Biological Roles of Resolvins and Related Substances in the Resolution of Pain". BioMed Research International. 2015: 830930. doi:10.1155/2015/830930. PMC 4538417. PMID 26339646.
- 1 2 Markworth JF, Maddipati KR, Cameron-Smith D (2016). "Emerging roles of pro-resolving lipid mediators in immunological and adaptive responses to exercise-induced muscle injury". Exercise Immunology Review. 22: 110–34. PMID 26853678.
- ↑ Serhan, C. N.; Arita, M; Hong, S; Gotlinger, K (2004). "Resolvins, docosatrienes, and neuroprotectins, novel omega-3-derived mediators, and their endogenous aspirin-triggered epimers". Lipids. 39 (11): 1125–32. doi:10.1007/s11745-004-1339-7. PMID 15726828.
- ↑ Chiang, N; Shinohara, M; Dalli, J; Mirakaj, V; Kibi, M; Choi, A. M.; Serhan, C. N. (2013). "Inhaled carbon monoxide accelerates resolution of inflammation via unique proresolving mediator-heme oxygenase-1 circuits". The Journal of Immunology. 190 (12): 6378–88. doi:10.4049/jimmunol.1202969. PMC 3679316. PMID 23650615.
- 1 2 Russell CD, Schwarze J (2014). "The role of pro-resolution lipid mediators in infectious disease". Immunology. 141 (2): 166–73. doi:10.1111/imm.12206. PMC 3904237. PMID 24400794.
- ↑ Zhang MJ, Sansbury BE, Hellmann J, Baker JF, Guo L, Parmer CM, Prenner JC, Conklin DJ, Bhatnagar A, Creager MA, Spite M (2016). "Resolvin D2 Enhances Postischemic Revascularization While Resolving Inflammation". Circulation. 134 (9): 666–80. doi:10.1161/CIRCULATIONAHA.116.021894. PMID 27507404.
- ↑ Skarke, C; Alamuddin, N; Lawson, J. A.; Li, X; Ferguson, J. F.; Reilly, M. P.; Fitzgerald, G. A. (2015). "Bioactive products formed in humans from fish oils". Journal of Lipid Research. 56 (9): 1808–20. doi:10.1194/jlr.M060392. PMC 4548785. PMID 26180051.
- ↑ Murphy, Robert C. (Sep 2015). "Specialized pro-resolving mediators: do they circulate in plasma?". J Lipid Res. 56 (9): 1641–2. doi:10.1194/jlr.C062356. PMC 4548769. PMID 26232500. Retrieved 27 September 2015.
- ↑ Dalli, Jesmond; Serhan, Charles N. (2012-10-11). "Specific lipid mediator signatures of human phagocytes: microparticles stimulate macrophage efferocytosis and pro-resolving mediators". Blood. 120 (15): e60–e72. doi:10.1182/blood-2012-04-423525. ISSN 0006-4971. PMC 3471524. PMID 22904297.
- ↑ Colas, Romain A.; Shinohara, Masakazu; Dalli, Jesmond; Chiang, Nan; Serhan, Charles N. (2014-07-01). "Identification and signature profiles for pro-resolving and inflammatory lipid mediators in human tissue". American Journal of Physiology. Cell Physiology. 307 (1): C39–C54. doi:10.1152/ajpcell.00024.2014. ISSN 0363-6143. PMC 4080182. PMID 24696140.
- ↑ Sasaki, Ai; Fukuda, Hayato; Shiida, Narumi; Tanaka, Nobuaki; Furugen, Ayako; Ogura, Jiro; Shuto, Satoshi; Mano, Nariyasu; Yamaguchi, Hiroaki (2015-01-11). "Determination of ω-6 and ω-3 PUFA metabolites in human urine samples using UPLC/MS/MS". Analytical and Bioanalytical Chemistry. 407 (6): 1625–1639. doi:10.1007/s00216-014-8412-5. ISSN 1618-2642. PMID 25577352.
- ↑ Keelan, Jeffrey A.; Mas, Emilie; D'Vaz, Nina; Dunstan, Janet A.; Li, Shaofu; Barden, Anne E.; Mark, Peter J.; Waddell, Brendan J.; Prescott, Susan L. (2015-02-01). "Effects of maternal n-3 fatty acid supplementation on placental cytokines, pro-resolving lipid mediators and their precursors". Reproduction. 149 (2): 171–178. doi:10.1530/REP-14-0549. ISSN 1470-1626. PMID 25504868.
- ↑ "A randomized controlled trial of the effects of n-3 fatty acids on resolvins in chronic kidney disease - Clinical Nutrition". www.clinicalnutritionjournal.com. Retrieved 2015-10-04.
- ↑ Wu, S. H.; Chen, X. Q.; Liu, B; Wu, H. J.; Dong, L (2013). "Efficacy and safety of 15(R/S)-methyl-lipoxin A(4) in topical treatment of infantile eczema". British Journal of Dermatology. 168 (1): 172–8. doi:10.1111/j.1365-2133.2012.11177.x. PMID 22834636.