Twin study
Twin studies reveal the importance of environmental and genetic influences for traits, phenotypes, and disorders. Twin research is considered a key tool in behavioral genetics and in content fields, from biology to psychology. Twin studies are part of the broader methodology used in behavior genetics, which uses any all data that are genetically informative – siblings studies, adoption studies, pedigree, etc. These studies have been used to track traits ranging from personal behavior to the presentation of severe mental illnesses such as schizophrenia.
Twins are a valuable source for observation because they allow the study of environmental influence and varying genetic makeup: "identical" or monozygotic (MZ) twins share nearly 100% of their genes, which means that most differences between the twins (such as height, susceptibility to boredom, intelligence, depression, etc.) is due to experiences that one twin has but not the other twin.[1] "Fraternal" or dizygotic (DZ) twins share only about 50% of their genes, the same as any other sibling. Twins also share many aspects of their environment (e.g., uterine environment, parenting style, education, wealth, culture, community) because they are born into the same family. The presence of a given genetic trait in only one member of a pair of identical twins (called discordance) provides a powerful window into environmental effects.
Twins are also useful in showing the importance of the unique environment (specific to one twin or the other) when studying trait presentation. Changes in the unique environment can stem from an event or occurrence that has only affected one twin. This could range form a head injury or a birth defect that one twin has sustained while the other remains healthy.
The classical twin design compares the similarity of monozygotic (identical) and dizygotic (fraternal) twins. If identical twins are considerably more similar than fraternal twins (which is found for most traits), this implicates that genes play an important role in these traits. By comparing many hundreds of families with twins, researchers can then understand more about the roles of genetic effects, shared environment, and unique environment in shaping behavior.
Modern twin studies have shown that almost all traits are in part influenced by genetic differences, with some characteristics showing a stronger influence (e.g. height), others an intermediate level (e.g. personality traits) and some more complex heritabilities, with evidence for different genes affecting different aspects of the trait — as in the case of autism.[2]
History

Twins have been of interest to scholars since early civilization, including the early physician Hippocrates (5th century BCE), who attributed similar diseases in twins to shared material circumstances, and the stoic philosopher Posidonius (1st century BCE), who attributed such similarities to shared astrological circumstances.[3] More recent study is from Sir Francis Galton's pioneering use of twins to study the role of genes and environment on human development and behavior. Galton, however, was unaware of the difference between identical and DZ twins.[4]
This factor was still not understood when the first study using psychological tests was conducted by Edward Thorndike (1905) using fifty pairs of twins. This paper was an early statement of the hypothesis that family effects decline with age. His study compared twin pairs age 9-10 and 13-14 to normal siblings born within a few years of one another.
Thorndike incorrectly reasoned that his data supported for there being one, not two, twin types. This mistake was repeated by Ronald Fisher (1919), who argued
- "The preponderance of twins of like sex, does indeed become a new problem, because it has been formerly believed to be due to the proportion of identical twins. So far as I am aware, however, no attempt has been made to show that twins are sufficiently alike to be regarded as identical really exist in sufficient numbers to explain the proportion of twins of like sex."[5]
An early, and perhaps first, study understanding the distinction is from the German geneticist Hermann Werner Siemens in 1924.[6] Chief among Siemens' innovations was the polysymptomatic similarity diagnosis. This allowed him to account for the oversight that had stumped Fisher, and was a staple in twin research prior to the advent of molecular markers.
Wilhelm Weinberg and colleagues in 1910 used the identical-DZ distinction to calculate respective rates from the ratios of same- and opposite-sex twins in a maternity population. They partitioned co-variation amongst relatives into genetic and environmental elements, anticipating the later work of Fisher and Wright, including the effect of dominance on similarity of relatives, and beginning the first classic-twin studies.[7]
Methods
The power of twin designs arises from the fact that twins may be either monozygotic (identical (MZ): developing from a single fertilized egg and therefore sharing all of their alleles) – or dizygotic (DZ: developing from two fertilized eggs and therefore sharing on average 50% of their polymorphic alleles, the same level of genetic similarity as found in non-twin siblings). These known differences in genetic similarity, together with a testable assumption of equal environments for identical and fraternal twins[8] creates the basis for the twin design for exploring the effects of genetic and environmental variance on a phenotype.[9][10]
The basic logic of the twin study can be understood with very little mathematics beyond an understanding of correlation and the concept of variance.
Like all behavior genetic research, the classic twin study begins from assessing the variance of a behavior (called a phenotype by geneticists) in a large group, and attempts to estimate how much of this is due to:
- genetic effects (heritability);
- shared environment – events that happen to both twins, affecting them in the same way;
- unshared, or unique, environment – events that occur to one twin but not the other, or events that affect either twin in a different way.
Typically these three components are called A (additive genetics) C (common environment) and E (unique environment); hence the acronym ACE. It is also possible to examine non-additive genetics effects (often denoted D for dominance (ADE model); see below for more complex twin designs).
The ACE model indicates what proportion of variance in a trait is heritable, versus the proportion due to shared environment or un-shared environment. Research is carried out using SEM programs such as OpenMx, however the core logic of the twin design is the same, as described below:
Monozygotic (identical – MZ) twins raised in a family share both 100% of their genes, and all of the shared environment. Any differences arising between them in these circumstances are random (unique). The correlation between identical twins provides an estimate of A + C. Dizygotic (DZ) twins also share C, but share on average 50% of their genes: so the correlation between fraternal twins is a direct estimate of ½A+C. If r is correlation, then rmz and rdz are simply the correlations of the trait in identical and fraternal twins respectively. For any particular trait, then:
- rmz = A + C
- rdz = ½A + C
A, therefore, is twice the difference between identical and fraternal twin correlations : the additive genetic effect (Falconer's formula). C is simply the MZ correlation minus this estimate of A. The random (unique) factor E is 1 − rmz: i.e., MZ twins differ due to unique environments only. (Jinks & Fulker, 1970; Plomin, DeFries, McClearn, & McGuffin, 2001).
Stated again, the difference between these two sums, then, allows us to solve for A, C, and E. As the difference between the identical and fraternal correlations is due entirely to a halving of the genetic similarity, the additive genetic effect 'A' is simply twice the difference between the identical and fraternal correlations:
- A = 2 (rmz − rdz)
As the identical correlation reflects the full effect of A and C, E can be estimated by subtracting this correlation from 1
- E = 1 − rmz
Finally, C can be derived:
- C = rmz − A
Modern modeling
Beginning in the 1970s, research transitioned to modeling genetic, environmental effects using maximum likelihood methods (Martin & Eaves, 1977). While computationally much more complex, this approach has numerous benefits rendering it almost universal in current research.
An example structural model (for the heritability of height among Danish males)[11] is shown:
 A: ACE model showing raw (non-standardised) variance coefficients |
 B: ACE model showing standardised variance coefficients |
Model A on the left shows the raw variance in height. This is useful as it preserves the absolute effects of genes and environments, and expresses these in natural units, such as mm of height change. Sometimes it is helpful to standardize the parameters, so each is expressed as percentage of total variance. Because we have decomposed variance into A, C, and E, the total variance is simply A + C + E. We can then scale each of the single parameters as a proportion of this total, i.e., Standardised–A = A/(A + C + E). Heritability is the standardised genetic effect.
Model comparison
A principal benefit of modeling is the ability to explicitly compare models: Rather than simply returning a value for each component, the modeler can compute confidence intervals on parameters, but, crucially, can drop and add paths and test the effect via statistics such as the AIC. Thus, for instance to test for predicted effects of family or shared environment on behavior, an AE model can be objectively compared to a full ACE model. For example, we can ask of the figure above for height: Can C (shared environment) be dropped without significant loss of fit? Alternatively, confidence intervals can be calculated for each path.
Multi-group and multivariate modeling
Multivariate modeling can give answers to questions about the genetic relationship between variables that appear independent. For instance: do IQ and long-term memory share genes? Do they share environmental causes? Additional benefits include the ability to deal with interval, threshold, and continuous data, retaining full information from data with missing values, integrating the latent modeling with measured variables, be they measured environments, or, now, measured molecular genetic markers such as SNPs. In addition, models avoid constraint problems in the crude correlation method: all parameters will lie, as they should, between 0–1 (standardized).
Multivariate, and multiple-time wave studies, with measured environment and repeated measures of potentially causal behaviours are now the norm. Examples of these models include extended twin designs,[12][13] simplex models,[14] and growth-curve models.[15]
SEM programs such as OpenMx[16] and other applications suited to constraints and multiple groups have made the new techniques accessible to reasonably skilled users.
Modeling the environment: MZ discordant designs
As MZ twins share both their genes and their family-level environmental factors, any differences between MZ twins reflect E: the unique environment. Researchers can use this information to understand the environment in powerful ways, allowing epidemiological tests of causality that are otherwise typically confounded by factors such as gene-environment covariance, reverse causation and confounding.
An example of a positive MZ discordant effect is shown below on the left. The twin who scores higher on trait 1 also scores higher on trait 2. This is compatible with a "dose" of trait 1 causing an increase in trait 2. Of course, trait 2 might also be affecting trait 1. Disentangling these two possibilities requires a different design (see below for an example). A null result is incompatible with a causal hypothesis.
 A depiction of MZ-discordance data |
 MZ discordant test of hypothesis that exercise protects against depression |
Take for instance the case of an observed link between depression and exercise (See Figure above on right). People who are depressed also reporting doing little physical activity. One might hypothesise that this is a causal link: that "dosing" patients with exercise would raise their mood and protect against depression. The next figure shows what empirical tests of this hypothesis have found: a null result.[17]
Longitudinal discordance designs
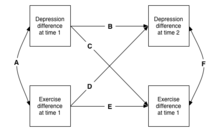
As may be seen in the next Figure, this design can be extended to multiple measurements, with consequent increase in the kinds of information that one can learn. This is called a cross-lagged model (multiple traits measured over more than one time).[18]
In the longitudinal discordance model, differences between identical twins can be used to take account of relationships among differences across traits at time one (path A), and then examine the distinct hypotheses that increments in trait1 drive subsequent change in that trait in the future (paths B and E), or, importantly, in other traits (paths C & D). In the example, the hypothesis that the observed correlation where depressed persons often also exercise less than average is causal, can be tested. If exercise is protective against depression, then path D should be significant, with a twin who exercises more showing less depression as a consequence.
Assumptions
It can be seen from the modeling above, the main assumption of the twin study is that of equal environments. This assumption has been directly tested. A special case occurs where parents believe their twins to be non-identical when in fact they are genetically identical. Studies of a range of psychological traits indicate that these children remain as concordant as MZ twins raised by parents who treated them as identical.[19]
Molecular genetic methods of heritability estimation have offered evidence that the equal environments assumption of the classic twin design may be sound.[20]
Measured similarity: A direct test of assumptions in twin designs
A particularly powerful technique for testing the twin method was reported by Visscher et al.[21] Instead of using twins, this group took advantage of the fact that while siblings on average share 50% of their genes, the actual gene-sharing for individual sibling pairs varies around this value, essentially creating a continuum of genetic similarity or "twinness" within families. Estimates of heritability based on direct estimates of gene sharing confirm those from the twin method, providing support for the assumptions of the method.
Sex differences
Genetic factors may differ between the sexes, both in gene expression and in the range of gene × environment interactions. Fraternal opposite sex twin pairs are invaluable in explicating these effects.
In an extreme case, a gene may only be expressed in one sex (qualitative sex limitation). More commonly, the effects of gene-alleles may depend on the sex of the individual. A gene might cause a change of 100 g in weight in males, but perhaps 150 g in females – a quantitative gene effect. Such effects are Environments may impact on the ability of genes to express themselves and may do this via sex differences. For instance genes affecting voting behavior would have no effect in females if females are excluded from the vote. More generally, the logic of sex-difference testing can extend to any defined sub-group of individuals. In cases such as these, the correlation for same and opposite sex DZ twins will differ, betraying the effect of the sex difference.
For this reason, it is normal to distinguish three types of fraternal twins. A standard analytic workflow would involve testing for sex-limitation by fitting models to five groups, identical male, identical female, fraternal male, fraternal female, and fraternal opposite sex. Twin modeling thus goes beyond correlation to test causal models involving potential causal variables, such as sex.
Gene × environment interactions
Gene effects may often be dependent on the environment. Such interactions are known as G×E interactions, in which the effects of a gene allele differ across different environments. Simple examples would include situations where a gene multiplies the effect of an environment: perhaps adding 1 inch to height in high nutrient environments, but only half an inch to height in low-nutrient environments. This is seen in different slopes of response to an environment for different genotypes.
Often researchers are interested in changes in heritability under different conditions: In environments where alleles can drive large phenotypic effects (as above), the relative role of genes will increase, corresponding to higher heritability in these environments.
A second effect is G × E correlation, in which certain alleles tend to accompany certain environments. If a gene causes a parent to enjoy reading, then children inheriting this allele are likely to be raised in households with books due to GE correlation: one or both of their parents has the allele and therefore will accumulate a book collection and pass on the book-reading allele. Such effects can be tested by measuring the purported environmental correlate (in this case books in the home) directly.
Often the role of environment seems maximal very early in life, and decreases rapidly after compulsory education begins. This is observed for instance in reading[22] as well as intelligence.[23] This is an example of a G*Age effect and allows an examination of both GE correlations due to parental environments (these are broken up with time), and of G*E correlations caused by individuals actively seeking certain environments.[24]
Norms of reaction
Studies in plants or in animal breeding allow the effects of experimentally randomized genotypes and environment combinations to be measured. By contrast, human studies are typically observational.[25][26] This may suggest that norms of reaction cannot be evaluated.[27][28]
As in other fields such as economics and epidemiology, several designs have been developed to capitalise on the ability to use differential gene-sharing, repeated exposures, and measured exposure to environments (such as children social status, chaos in the family, availability and quality of education, nutrition, toxins etc.) to combat this confounding of causes. An inherent appeal of the classic twin design is that it begins to untangle these confounds. For example, in identical and fraternal twins shared environment and genetic effects are not confounded, as they are in non-twin familial studies.[10] Twin studies are thus in part motivated by an attempt to take advantage of the random assortment of genes between members of a family to help understand these correlations.
While the twin study tells us only how genes and families affect behavior within the observed range of environments, and with the caveat that often genes and environments will covary, this is a considerable advance over the alternative, which is no knowledge of the different roles of genes and environment whatsoever.[29] Twin studies are therefore often used as a method of controlling at least one part of this observed variance: Partitioning, for instance, what might previously have been assumed to be family environment into shared environment and additive genetics using the experiment of fully and partly shared genomes in twins.[29]
No single design can address all issues. Additional information is available outside the classic twin design. Adoption designs are a form of natural experiment that tests norms of reaction by placing the same genotype in different environments.[30] Association studies, e.g.,[31] allow direct study of allelic effects. Mendelian randomization of alleles also provides opportunities to study the effects of alleles at random with respect to their associated environments and other genes.[32]
Extended twin designs and more complex genetic models
The basic or classical twin-design contains only identical and fraternal twins raised in their biological family. This represents only a sub-set of the possible genetic and environmental relationships. It is fair to say, therefore, that the heritability estimates from twin designs represent a first step in understanding the genetics of behavior.
The variance partitioning of the twin study into additive genetic, shared, and unshared environment is a first approximation to a complete analysis taking into account gene-environment covariance and interaction, as well as other non-additive effects on behavior. The revolution in molecular genetics has provided more effective tools for describing the genome, and many researchers are pursuing molecular genetics in order to directly assess the influence of alleles and environments on traits.
An initial limitation of the twin design is that it does not afford an opportunity to consider both Shared Environment and Non-additive genetic effects simultaneously. This limit can be addressed by including additional siblings to the design.
A second limitation is that gene-environment correlation is not detectable as a distinct effect. Addressing this limit requires incorporating adoption models, or children-of-twins designs, to assess family influences uncorrelated with shared genetic effects.
Continuous variables and ordinal variables
While concordance studies compare traits either present or absent in each twin, correlational studies compare the agreement in continuously varying traits across twins.
Criticism
The Twin Method has been subject to criticism from statistical genetics, statistics, and psychology, with some arguing that conclusions reached via this method are ambiguous or meaningless. Core elements of these criticisms and their rejoinders are listed below.
Criticisms of statistical methods
It has been argued that the statistical underpinnings of twin research are invalid. Such statistical critiques argue that heritability estimates used for most twin studies rest on restrictive assumptions that are usually not tested, and if they are, can often found violated by the data.
For example, Peter Schonemann has criticized methods for estimating heritability developed in the 1970s. He has also argued that the heritability estimate from a twin study may reflect factors other than shared genes. Using the statistical models published in Loehlin and Nichols (1976),[33] the narrow heritability’s of HR of responses to the question “did you have your back rubbed” has been shown to work out to .92 heritable for males and .21 heritable for females, and the question “Did you wear sunglasses after dark?” is 130% heritable for males and 103% for females[34][35]
Responses to statistical critiques
Before computers, statisticians used methods that were computationally tractable, at the cost of known limitations. Since the 1980s these approximate statistical methods have been discarded: Modern twin methods based on structural equation modeling are not subject to the limitations and heritability estimates such as those noted above are mathematically impossible.[36] Critically, the newer methods allow for explicit testing of the role of different pathways and incorporation and testing of complex effects.[29]
Sampling: Twins as representative members of the population
Results of twin studies cannot be automatically generalized beyond the population they come from. It is therefore important to understand the particular sample studied, and the nature of twins themselves. Twins are not a random sample of the population, and they differ in their developmental environment. In this sense they are not representative.[37]
For example: Dizygotic (DZ) twin births are affected by many factors. Some women frequently produce more than one egg at each menstrual period and, therefore, are more likely to have twins. This tendency may run in the family either in the mother's or father's side of the family, and often runs through both. Women over the age of 35 are more likely to produce two eggs. Women who have three or more children are also likely to have dizygotic twins. Artificial induction of ovulation and in vitro fertilization-embryo replacement can also give rise to fraternal and identical twins.[38][39][40][41][42][43]
Response to representativeness of twins
Twins differ very little from non-twin siblings. Measured studies on the personality and intelligence of twins suggest that they have scores on these traits very similar to those of non-twins (for instance Deary et al. 2006).
Separated twin pairs as representative of other twins
Separated twin pairs, identical or fraternal, are generally separated by adoption. This makes their families of origin non-representative of typical twin families in that they adopt their children away. The families they are adopted to are also non-representative of typical twin families in that they are all approved for adoption by children's protection authorities and that a disproportionally large fraction of them have no biological children. Those who volunteer to studies are not even representative of separated twins in general since not all separated twins agree to be part of twin studies.[44][45]
Detection problems
There can be some issues of undetected behaviors in the case of behaviors that many people keep secret presently or in their earlier lives. They may not be as willing to reveal behaviors that are discriminated against or stigmatized. If environment played no role in the actual behavior, skewed detection would still make it look like it played a role. For environment to appear to have no role in such cases, there would have to be either a counterproductivity of intolerance in the sense of intolerance causing the behavior it is bigoted against, or a flaw in the study that makes the results scientifically useless. Even if environment does play a role, the numbers would still be skewed.[46][47][48]
Physical few-character stereotypes causing behavioral correlations
Apparent correlation between heredity and behavior may be due to social stereotypes making people with certain physical characteristics (which may be hereditary physical characteristics) behave in certain ways, without any biological link between the genetic factor and the behavior. This violatates the equal environment assumption of twin design and suggests a stronger argument for environmental influences.[49] This factor has an especially high likelihood of being the case for behaviors that are culturally depicted in mass media as being more accepted by those who consider them to be biological than by those who consider them learned (e.g. sexuality and some psychiatric diagnoses) and for physical characteristics that are not culturally considered by mass media to be bigoted to associate with certain behaviors (e.g. not ethnic or racial). Such unequal environment biases may concentrate on a relatively small number of minor physical characteristics that are subject to stereotypes that are considered to be politically correct, with no statistically significant effect on overall ratings of physical similarity, let alone anywhere near being able to pass identical twins off as being fraternal.[50][51][52]
Terminology
Pairwise concordance
For a group of twins, pairwise concordance is defined as C/(C+D), where C is the number of concordant pairs and D is the number of discordant pairs.
For example, a group of 10 twins have been pre-selected to have one affected member (of the pair). During the course of the study four other previously non-affected members become affected, giving a pairwise concordance of 4/(4+6) or 4/10 or 40%.
Probandwise concordance
For a group of twins in which at least one member of each pair is affected, probandwise concordance is a measure of the proportion of twins who have the illness who have an affected twin and can be calculated with the formula of 2C/(2C+D), in which C is the number of concordant pairs and D is the number of discordant pairs.
For example, consider a group of 10 twins that have been pre-selected to have one affected member. During the course of the study, four other previously non-affected members become affected, giving a probandwise concordance of 8/(8+6) or 8/14 or 57%.
See also
- Genome-wide complex trait analysis
- Minnesota Twin Family Study
- Behavioral genetics
- "Burt Affair"
- Gene-environment interaction
- Gene-environment correlation
- Heritability
- Heritability of IQ
- Human nature
- Identical Strangers: A Memoir of Twins Separated and Reunited
- Kaiser Wilhelm Institute of Anthropology, Human Heredity, and Eugenics
- Michigan State University Twin Registry
- Nature versus nurture
- Otmar Freiherr von Verschuer
- Quantitative genetics
- Differential susceptibility
- TwinsUk
References
- ↑ Plomin, R.; DeFries, J. C.; Knopik, V. S.; Neiderhiser, J. M. (Ed.). (2014). Behavioral Genetics (6th ed.). New York, NY: Worth Publishers
- ↑ Polderman, Tinca J C; Benyamin, Beben; de Leeuw, Christiaan A; Sullivan, Patrick F; van Bochoven, Arjen; Visscher, Peter M; Posthuma, Danielle (2015). "Meta-analysis of the heritability of human traits based on fifty years of twin studies". Nature Genetics. 47 (7): 702–709. doi:10.1038/ng.3285. ISSN 1061-4036.
- ↑ Cicero, De Divinatione, (On Divination), ii. 42
- ↑ Rende, R. D.; Plomin, R.; Vandenberg, S. G. (1990). "Who discovered the twin method?". Behavior Genetics. 20 (2): 277–285. doi:10.1007/BF01067795. PMID 2191648.
- ↑ Fisher, R. A. (1919). "The Genesis of Twins". Genetics. 4 (5): 489–499. PMC 1200469
 . PMID 17245935.
. PMID 17245935. - ↑ Siemens, Hermann Werner (1924). Die zwillingspathologie; ihre bedeutung, ihre methodik, ihre bisherigen ergebnisse. Berlin: Springer. OCLC 18362377.
- ↑ Crow, James F. (1999). "Hardy, Weinberg and language impediments". Genetics. 152 (3): 821–825. PMC 1460671. PMID 10388804.
- ↑ Propping, Peter; Bouchard, Thomas J., eds. (1993). Twins as a tool of behavioral genetics. London: J. Wiley. p. 326. ISBN 0-471-94174-3.
- ↑ Cardon, Lon R.; Neale, Michael C. (1992). Methodology for genetic studies of twins and families. Boston: Kluwer Academic Publishers. ISBN 0-7923-1874-9.
- 1 2 Martin, Nicholas; Boomsma, Dorret; Machin, Geoffrey (1997). "A twin-pronged attack on complex traits" (PDF). Nature Genetics. 17 (4): 387–392. doi:10.1038/ng1297-387. PMID 9398838. hdl:1871/2733.
- ↑ Silventoinen, K.; Sammalisto, S.; Perola, M.; Boomsma, D. I.; Cornes, B. K.; Davis, C.; Dunkel, L.; De Lange, M.; Harris, J. R.; Hjelmborg, J. V. B.; Luciano, M.; Martin, N. G.; Mortensen, J.; Nisticò, L.; Pedersen, N. L.; Skytthe, A.; Spector, T. D.; Stazi, M. A.; Willemsen, G.; Kaprio, J. (October 2003). "Heritability of Adult Body Height: A Comparative Study of Twin Cohorts in Eight Countries". Twin Research. 6 (5): 399–408. CiteSeerX 10.1.1.81.3898. doi:10.1375/136905203770326402. PMID 14624724.
- ↑ Keller, M. C.; Medland, S. E.; Duncan, L. E. (May 2010). "Are Extended Twin Family Designs Worth the Trouble? A Comparison of the Bias, Precision, and Accuracy of Parameters Estimated in Four Twin Family Models". Behavior Genetics. 40 (3): 377–393. doi:10.1007/s10519-009-9320-x. PMC 3228846. PMID 20013306.
- ↑ Coventry, W. L.; Keller, M. C. (June 2005). "Estimating the Extent of Parameter Bias in the Classical Twin Design: A Comparison of Parameter Estimates from Extended Twin-Family and Classical Twin Designs". Twin Research and Human Genetics. 8 (3): 214–223. doi:10.1375/1832427054253121. PMID 15989749.
- ↑ Gillespie, Nathan A.; Evans, David E.; Wright, Margie. M.; Martin, Nicholas G. (2004). "Genetic Simplex Modeling of Eysenck's Dimensions of Personality in a Sample of Young Australian Twins" (PDF). Twin Research. 7 (6): 637–648. doi:10.1375/1369052042663814. PMID 15607015.
- ↑ Neale, M. C.; McArdle, J. J. (September 2000). "Structured latent growth curves for twin data" (PDF). Twin Research. 3 (3): 165–177. doi:10.1375/136905200320565454. PMID 11035490.
- ↑ Boker, Steven; Neale, Michael; Maes, Hermine; Wilde, Michael; Spiegel, Michael; Brick, Timothy; Spies, Jeffrey; Estabrook, Ryne; Kenny, Sarah; Bates, Timothy; Mehta, Paras; Fox, John (2011). "OpenMx: An Open Source Extended Structural Equation Modeling Framework". Psychometrika. 76 (2): 306–317. doi:10.1007/s11336-010-9200-6.
- ↑ M. H. De Moor, D. I. Boomsma, J. H. Stubbe, G. Willemsen and E. J. de Geus. (2008). Testing causality in the association between regular exercise and symptoms of anxiety and depression. Archives of general psychiatry, 65, 897-905. doi
- ↑ S. A. Burt, M. McGue and W. G. Iacono. (2009). Nonshared environmental mediation of the association between deviant peer affiliation and adolescent externalizing behaviors over time: results from a cross-lagged monozygotic twin differences design. Dev Psychol, 45, 1752–60. doi
- ↑ Kendler, K.S., Neale, M.C., Kessler, R.C., Heath, A.C. and Eaves, L.J., 1993. Panic disorder in women: A population-based twin study. Psychological Medicine 23, pp. 397–406
- ↑ Turkheimer, E. (2011). "Still Missing". Research in Human Development. 8 (3–4): 227–241. doi:10.1080/15427609.2011.625321.
- ↑ Visscher, Peter M.; Medland, Sarah E.; Ferreira, Manuel A. R.; Morley, Katherine I.; Zhu, Gu; Cornes, Belinda K.; Montgomery, Grant W.; Martin, Nicholas G. (2006). "Assumption-Free Estimation of Heritability from Genome-Wide Identity-by-Descent Sharing between Full Siblings". PLoS Genetics. 2 (3): e41. doi:10.1371/journal.pgen.0020041. PMC 1413498. PMID 16565746.
- ↑ Byrne, Brian; Wadsworth, Sally; Corley, Robin; Samuelsson, Stefan; Quain, Peter; Defries, John C.; Willcutt, Erik; Olson, Richard K. (2005). "Longitudinal Twin Study of Early Literacy Development: Preschool and Kindergarten Phases" (PDF). Scientific Studies of Reading. 9 (3): 219–235. doi:10.1207/s1532799xssr0903_3.
- ↑ Deary, Ian J.; Spinath, Frank M.; Bates, Timothy C. (2006). "Genetics of intelligence". European Journal of Human Genetics. 14 (6): 690–700. doi:10.1038/Sj.Ejhg.5201588. PMID 16721405.
- ↑ Plomin, Robert; Daniels, Denise (1987). "Why are children in the same family so different from one another?". Behavioral and Brain Sciences. 10: 1–16. doi:10.1017/S0140525X00055941.
revisited in: Plomin, R.; Daniels, D. (June 2011). "Why are children in the same family so different from one another?". International Journal of Epidemiology. 40 (3): 563–582. doi:10.1093/ije/dyq148. PMC 3147063. PMID 21807642. - ↑ Kempthorne, Oscar (1997). "Heritability: uses and abuses". Genetica. 99 (2-3): 109–112. doi:10.1023/A:1018352202363. PMID 9463066.
- ↑ Kendler, K. S.; Gruenberg, A. M. (June 1984). "An Independent Analysis of the Danish Adoption Study of Schizophrenia: VI. The Relationship Between Psychiatric Disorders as Defined by DSM-III in the Relatives and Adoptees". Archives of General Psychiatry. 41 (6): 555–564. doi:10.1001/archpsyc.1984.01790170029004. PMID 6732417.
- ↑ Kamin, Leon J.; Rose, Steven R.; Lewontin, Richard C. (1984). Not in Our Genes: Biology, Ideology and Human Nature. New York: Penguin Books. ISBN 0-14-022605-2.
- ↑ Rose, Richard J. (1982). "Separated Twins: Data and Their Limits". Science. 215 (4535): 959–960. Bibcode:1982Sci...215..959F. doi:10.1126/science.215.4535.959. PMID 17821364.
- 1 2 3 M. C. Neale and H. H. Maes. (1996). Methodology for genetics studies of twins and families. Journal.
- ↑ Petrill, S. A.; Deater-Deckard, K. (July–August 2004). "The heritability of general cognitive ability: A within-family adoption design". Intelligence. 32 (4): 403–201. doi:10.1016/j.intell.2004.05.001.
- ↑ Steer, C. D.; Davey Smith, G.; Emmett, P. M.; Hibbeln, J. R.; Golding, J. (July 2010). Penha-Goncalves, Carlos, ed. "FADS2 Polymorphisms Modify the Effect of Breastfeeding on Child IQ". PLoS ONE. 5 (7): e11570. doi:10.1371/journal.pone.0011570. PMC 2903485. PMID 20644632.
- ↑ e.g. Davey Smith, G. (September 2007). "Capitalizing on Mendelian randomization to assess the effects of treatments". Journal of the Royal Society of Medicine. 100 (9): 432–435. doi:10.1258/jrsm.100.9.432. PMC 1963388. PMID 17766918.
- ↑ Loehlin, John C.; Nichols, Robert (1976). Heredity, environment, & personality: a study of 850 sets of twins. Austin: University of Texas Press. ISBN 0-292-73003-9. JSTOR 2826060.
- ↑ Schönemann, Peter H. (1997). "On models and muddles of heritability". Genetica. 99 (2–3): 97–108. doi:10.1023/A:1018358504373. PMID 9463078.
- ↑ Schönemann, Peter H. (1995). Totems of the IQ Myth: General Ability (g) and its Heritabilities (h2, HR). 1995 Meetings of the American Association for the Advancement of Sciences.
- ↑ M. C. Neale, S. M. Boker, G. Xie and H. H. Maes. (2002). Mx: Statistical Modelling. Journal.
- ↑ Record, R. G.; McKeown, Thomas; Edwards, J. H. (1970). "An investigation of the difference in measured intelligence between twins and single births". Annals of Human Genetics. 34 (1): 11–20. doi:10.1111/j.1469-1809.1970.tb00215.x. PMID 5529232.
- ↑ Woollett, Anne; Clegg, Averil (1983). Twins: from conception to five years. London: Century. p. 127. ISBN 0-7126-0204-6. OCLC 12445470.
- ↑ Corson, S. L.; Dickey, R. P.; Gocial, B.; Batzer, F. R.; Eisenberg, E.; Huppert, L.; Maislin, G. (1989). "Outcome in 242 in vitro fertilization-embryo replacement or gamete intrafallopian transfer-induced pregnancies". Fertility and Sterility. 51 (4): 644–650. PMID 2924931.
- ↑ Derom, C.; Derom, R.; Vlietinck, R.; Berghe, H. V.; Thiery, M. (May 1987). "Increased Monozygotic Twinning Rate After Ovulation Induction". The Lancet. 329 (8544): 1236–1238. doi:10.1016/S0140-6736(87)92688-2. PMID 2884372.
- ↑ Edwards, R. G.; Mettler, L.; Walters, D. E. (April 1986). "Identical twins and in vitro fertilization". Journal of In Vitro Fertilization and Embryo Transfer. 3 (2): 114–117. doi:10.1007/BF01139357. PMID 3701181.
- ↑ Leigh, Gillian (1983). All About Twins: A Handbook for Parents. London: Routledge and Kegan Paul. ISBN 978-0-7100-9888-7.
- ↑ Capron, Christiane; Vetta, Adrian R.; Duyme, Michel; Vetta, Atam (1999). "Misconceptions of biometrical IQists". Cahiers de Psychologie Cognitive/Current Psychology of Cognition. 18 (2): 115–160.
- ↑ Fatal Flaws in the Twin Study Paradigm: A Reply to Hatemi and Verhulst, Doron Shultziner 2013
- ↑ Twin Studies of Political Behavior: Untenable Assumptions?, Jon Beckwith and Corey A. Morris 2008
- ↑ Critical Analysis: A Comparison of Critical Thinking Changes in Psychology and Philosophy Classes, Teaching of Psychology 2014 41: 28
- ↑ Association for Psychological Science: Why Science Is Not Necessarily Self-Correcting, John P. A. Ioannidis 2012
- ↑ How Black African and White British Women Perceive Depression and Help-Seeking: a Pilot Vignette Study, International Journal of Social Psychiatry March 2010
- ↑ Segal, Nancy L. (1999). Entwined Lives. United States of America: Penguin Group. p. 3. ISBN 0-525-94465-6.
For example, some people have argued that identical twins are so alike because theyare treated more alike than fraternal twins, violating the equal environment assumption of the twin design, and that their more similar genetic design has nothing to with it.
- ↑ http://www.studentsatthecenter.org/sites/scl.dl-dev.com/files/Mind%20Brain%20Education.pdf
- ↑ The Unpredictable Species: What Makes Humans Unique, Philip Lieberman 2013
- ↑ "Mind, Brain and Education, Kurt Fischer, Christina Hinton
Further reading
- Free courseware, textbook, software, and example scripts for twin research
- Jang, K.L., McCrae, R.R., Angleitner, A. Riemann, R. & Livesley, W.J. (1998). Heritability of facet-level traits in a cross-cultural twin sample: support for a hierarchical model of personality. Journal of Personality and Social Psychology 74:1556-1565.
- R. Plomin, J. C. DeFries, V. S. Knopik and J. M. Neiderhiser. (2012). Behavioral Genetics. Worth Publishers. London
- Nancy L. Segal (2005) Indivisible by Two: Lives of Extraordinary Twins. New York, Harvard University Press.
- Segal, Nancy L. (2012). Born Together—Reared Apart. Cambridge (MA): Harvard University Press. ISBN 978-0-674-05546-9. Lay summary (16 May 2013).
- Am J Med Genet C Semin Med Genet. 2009 May 15;151C(2):136-41. Not really identical: epigenetic differences in monozygotic twins and implications for twin studies in psychiatry. Haque FN, Gottesman II, Wong AH.
Critical accounts
- Peter Schönemann (1997). Models and muddles of heritability. Genetica, 99, 97-108
- Peter Schönemann and Roberta D. Schönemann (1994). Environmental versus genetic models for Osborne’s personality data on identical and fraternal twins. CPC, 13 (2), 141-167
- Kamin, L. J. (1974). The Science and Politics of I.Q. Potomac, MD: Lawrence Erlbaum Associates.
- Kempthorne O. (1997). Heritability: uses and abuses. Genetica, Volume 99, Numbers 2-3, 1997, pp. 109–112(4)
- Joseph, J. (2003). The Gene Illusion: Genetic Research in Psychiatry and Psychology Under the Microscope. PCCS Books.
- This book has been critically reviewed for the American Psychological Association. Hanson, D. R. (2005). 'The Gene Illusion Confusion: A review of The Gene Illusion: Genetic Research in Psychiatry and Psychology Under the Microscope by Jay Joseph' [Electronic Version]. PsycCritiques, 50, e14.
- Christiane Capron, Adrian R. Vetta, Michel Duyme and Atam Vetta (1999). Misconceptions of biometrical IQists. Cahiers de Psychologie Cognitive/Current Psychology of Cognition 1999, 18 (2), 115-160
- Horwitz AV, Videon TM, Schmitz MF, Davis D. Rethinking twins and environments: possible social sources for assumed genetic influences in twin research. J Health Soc Behav. 2003 Jun;44(2):111-129.
- And in reply to this article see:
- Freese J and Powell B Tilting at Twindmills: rethinking sociological responses to behavioral genetics. J Health Soc Behav. 2003 Jun;44(2):130-135.
External links
Academic bodies
Several academic bodies exist to support behavior genetic research, including the Behavior Genetics Association, the International Society for Twin Studies, and the International Behavioural and Neural Genetics Society. Behavior genetic work also features prominently in several more general societies, for instance the International Society of Psychiatric Genetics.
Journals
Prominent specialist journals in the field include Behavior Genetics, Genes, Brain and Behavior, and Twin Research and Human Genetics.