Butyric acid
| |||
 | |||
| Names | |||
|---|---|---|---|
| Preferred IUPAC name
Butanoic acid[1] | |||
| Other names | |||
| Identifiers | |||
| 107-92-6 | |||
| 3D model (Jmol) | Interactive image Interactive image | ||
| ChEBI | CHEBI:30772 | ||
| ChEMBL | ChEMBL14227 | ||
| ChemSpider | 259 | ||
| DrugBank | DB03568 | ||
| ECHA InfoCard | 100.003.212 | ||
| EC Number | 203-532-3 | ||
| 1059 | |||
| KEGG | C00246 | ||
| MeSH | Butyric+acid | ||
| PubChem | 264 | ||
| RTECS number | ES5425000 | ||
| UNII | 40UIR9Q29H | ||
| UN number | 2820 | ||
| |||
| |||
| Properties | |||
| C 3H 7COOH | |||
| Molar mass | 88.11 g·mol−1 | ||
| Appearance | Colorless liquid | ||
| Odor | Unpleasant and obnoxious | ||
| Density | 1.135 g/cm3 (−43 °C)[2] 0.9528 g/cm3 (25 °C)[3] | ||
| Melting point | −5.1 °C (22.8 °F; 268.0 K)[3] | ||
| Boiling point | 163.75 °C (326.75 °F; 436.90 K)[3] | ||
| Sublimes at −35 °C ΔsublH | |||
| Miscible | |||
| Solubility | Slightly soluble in CCl4[5] Miscible with ethanol, ether | ||
| log P | 0.79[5] | ||
| Vapor pressure | 0.112 kPa (20 °C)[6] 0.74 kPa (50 °C) 9.62 kPa (100 °C)[4] | ||
| Henry's law constant (kH) |
5.35·10−4 L·atm/mol[5] | ||
| Acidity (pKa) | 4.82[5] | ||
| Thermal conductivity | 1.46·105 W/m·K | ||
| Refractive index (nD) |
1.398 (20 °C)[3] | ||
| Viscosity | 1.814 cP (15 °C)[7] 1.426 cP (25 °C)[5] | ||
| Structure | |||
| Monoclinic (−43 °C)[2] | |||
| C2/m[2] | |||
| α = 90°, β = 111.45°, γ = 90° | |||
| 0.93 D (20 °C)[7] | |||
| Thermochemistry | |||
| 178.6 J/mol·K[4][5] | |||
| Std molar entropy (S |
222.2 J/mol·K[7] | ||
| Std enthalpy of formation (ΔfH |
−533.9 kJ/mol[4] | ||
| Std enthalpy of combustion (ΔcH |
2183.5 kJ/mol[4] | ||
| Hazards | |||
| Safety data sheet | External MSDS | ||
| GHS pictograms |  [8] [8] | ||
| GHS signal word | Danger | ||
| H314[8] | |||
| P280, P305+351+338, P310[8] | |||
| EU classification (DSD) |
| ||
| R-phrases | R20/21/22, R34, R36/37/38 | ||
| S-phrases | S26, S36, S45 | ||
| NFPA 704 | |||
| Flash point | 71 to 72 °C (160 to 162 °F; 344 to 345 K)[6][8] | ||
| 440 °C (824 °F; 713 K)[8] | |||
| Explosive limits | 2.2–13.4%[6] | ||
| Lethal dose or concentration (LD, LC): | |||
| LD50 (median dose) |
2000 mg/kg (oral, rat) | ||
| Related compounds | |||
| Other anions |
Butyrate | ||
| Related Carboxylic acids |
Propionic acid Acrylic acid Succinic acid Malic acid Tartaric acid Crotonic acid Fumaric acid Pentanoic acid | ||
| Related compounds |
1-Butanol Butyraldehyde Methyl butyrate | ||
| Except where otherwise noted, data are given for materials in their standard state (at 25 °C [77 °F], 100 kPa). | |||
| | |||
| Infobox references | |||


Butyric acid (from Greek βούτῡρον, meaning "butter"), also known under the systematic name butanoic acid, abbreviated BTA,[6] is a carboxylic acid with the structural formula CH3CH2CH2-COOH. Salts and esters of butyric acid are known as butyrates or butanoates. Butyric acid is found in milk, especially goat, sheep and buffalo milk, butter, parmesan cheese, and as a product of anaerobic fermentation (including in the colon and as body odor). It is also found in milk chocolate produced by the Hershey process, or added to imitate the flavour of Hershey's chocolate.[9] Butyric acid is present in, and is the main distinctive smell of, human vomit.[10] It has an unpleasant smell and acrid taste, with a sweetish aftertaste similar to ether. Mammals with good scent detection abilities, such as dogs, can detect it at 10 parts per billion, whereas humans can detect it in concentrations above 10 parts per million.
Butyric acid was first observed in impure form in 1814 by the French chemist Michel Eugène Chevreul. By 1818, he had purified it sufficiently to characterize it. However, Chevreul did not publish his early research on butyric acid; instead, he deposited his findings in manuscript form with the secretary of the Academy of Sciences in Paris, France. Henri Braconnot, a French chemist, was also researching the composition of butter and was publishing his findings, and this led to disputes about priority. As early as 1815, Chevreul claimed that he had found the substance responsible for the smell of butter.[11] By 1817, he published some of his findings regarding the properties of butyric acid and named it.[12] However, it was not until 1823 that he presented the properties of butyric acid in detail.[13] The name of butyric acid comes from the Latin word for butter, butyrum (or buturum), the substance in which butyric acid was first found.
Chemistry
Butyric acid is a fatty acid occurring in the form of esters in animal fats. The triglyceride of butyric acid makes up 3–4% of butter. When butter goes rancid, butyric acid is liberated from the glyceride by hydrolysis, leading to the unpleasant odor. It is an important member of the fatty acid subgroup called short-chain fatty acids. Butyric acid is a medium-strong acid that reacts with bases and strong oxidants, and attacks many metals.[14]
The acid is an oily, colorless liquid that is easily soluble in water, ethanol, and ether, and can be separated from an aqueous phase by saturation with salts such as calcium chloride. It is oxidized to carbon dioxide and acetic acid using potassium dichromate and sulfuric acid, while alkaline potassium permanganate oxidizes it to carbon dioxide. The calcium salt, Ca(C4H7O2)2·H2O, is less soluble in hot water than in cold.
Butyric acid has a structural isomer called isobutyric acid (2-methylpropanoic acid).
Safety
Personal protective equipment such as rubber or PVC gloves, protective eye goggles, and chemical-resistant clothing and shoes are used to minimize risks when handling butyric acid.
Inhalation of butyric acid may result in soreness of throat, coughing, a burning sensation, and laboured breathing. Ingestion of the acid may result in abdominal pain, shock, and collapse. Physical exposure to the acid may result in pain, blistering and skin burns, while exposure to the eyes may result in pain, severe deep burns and loss of vision.[14]
Production
It is industrially prepared by the fermentation of sugar or starch, brought about by the addition of putrefying cheese, with calcium carbonate added to neutralize the acids formed in the process. The butyric fermentation of starch is aided by the direct addition of Bacillus subtilis. Salts and esters of the acid are called butyrates or butanoates.
Butyric acid or fermentation butyric acid is also found as a hexyl ester hexyl butyrate in the oil of Heracleum giganteum (a type of hogweed) and as the octyl ester octyl butyrate in parsnip (Pastinaca sativa); it has also been noticed in skin flora and perspiration.
Uses
Butyric acid is used in the preparation of various butyrate esters. Low-molecular-weight esters of butyric acid, such as methyl butyrate, have mostly pleasant aromas or tastes. As a consequence, they are used as food and perfume additives. It is also used as an animal feed supplement due to the ability to reduce pathogenic bacterial colonization.[15] It is an approved food flavoring in the EU FLAVIS database (number 08.005).
Due to its powerful odor, it has also been used as a fishing bait additive.[16] Many of the commercially available flavors used in carp (Cyprinus carpio) baits use butyric acid as their ester base; however, it is not clear whether fish are attracted by the butyric acid itself or the substances added to it. Butyric acid was, however, one of the few organic acids shown to be palatable for both tench and bitterling.[17]
The substance has also been used as a stink bomb by Sea Shepherd Conservation Society to disrupt Japanese whaling crews,[18] as well as by anti-abortion protesters to disrupt abortion clinics.[19]
Biochemistry
Biosynthesis
Butyrate is produced as end-product of a fermentation process solely performed by obligate anaerobic bacteria. Fermented Kombucha "tea" includes butyric acid as a result of the fermentation. This fermentation pathway was discovered by Louis Pasteur in 1861. Examples of butyrate-producing species of bacteria:
- Clostridium butyricum
- Clostridium kluyveri
- Clostridium pasteurianum
- Faecalibacterium prausnitzii
- Fusobacterium nucleatum
- Butyrivibrio fibrisolvens
- Eubacterium limosum
The pathway starts with the glycolytic cleavage of glucose to two molecules of pyruvate, as happens in most organisms. Pyruvate is then oxidized into acetyl coenzyme A using a unique mechanism that involves an enzyme system called pyruvate:ferredoxin oxidoreductase. Two molecules of carbon dioxide (CO2) and two molecules of elemental hydrogen (H2) are formed as waste products from the cell. Then,
| Action | Responsible enzyme |
|---|---|
| Acetyl coenzyme A converts into acetoacetyl coenzyme A | acetyl-CoA-acetyl transferase |
| Acetoacetyl coenzyme A converts into β-hydroxybutyryl CoA | β-hydroxybutyryl-CoA dehydrogenase |
| β-hydroxybutyryl CoA converts into crotonyl CoA | crotonase |
| Crotonyl CoA converts into butyryl CoA (CH3CH2CH2C=O-CoA) | butyryl CoA dehydrogenase |
| A phosphate group replaces CoA to form butyryl phosphate | phosphobutyrylase |
| The phosphate group joins ADP to form ATP and butyrate | butyrate kinase |
ATP is produced, as can be seen, in the last step of the fermentation. Three molecules of ATP are produced for each glucose molecule, a relatively high yield. The balanced equation for this fermentation is
- C6H12O6 → C4H8O2 + 2 CO2 + 2 H2.
Several species form acetone and n-butanol in an alternative pathway, which starts as butyrate fermentation. Some of these species are:
- Clostridium acetobutylicum, the most prominent acetone and propianol producer, used also in industry
- Clostridium beijerinckii
- Clostridium tetanomorphum
- Clostridium aurantibutyricum
These bacteria begin with butyrate fermentation, as described above, but, when the pH drops below 5, they switch into butanol and acetone production to prevent further lowering of the pH. Two molecules of butanol are formed for each molecule of acetone.
The change in the pathway occurs after acetoacetyl CoA formation. This intermediate then takes two possible pathways:
- acetoacetyl CoA → acetoacetate → acetone
- acetoacetyl CoA → butyryl CoA → butyraldehyde → butanol
Highly-fermentable fiber residues, such as those from resistant starch, oat bran, pectin, and guar are transformed by colonic bacteria into short-chain fatty acids (SCFA) including butyrate, producing more SCFA than less fermentable fibers such as celluloses.[20] One study found that resistant starch consistently produces more butyrate than other types of dietary fiber.[21] The production of SCFA from fibers in ruminant animals such as cattle is responsible for the butyrate content of milk and butter.[22]
Fructans are another source of prebiotic soluble dietary fibers. They are often found in the soluble fibers of foods which are high in sulfur, such as the Allium and Cruciferous vegetables. Sources of fructans include wheat (although some wheat strains such as spelt contain lower amounts),[23] rye, barley, onion, garlic, Jerusalem and globe artichoke, asparagus, beetroot, chicory, dandelion leaves, leek, radicchio, the white part of spring onion, broccoli, brussels sprouts, cabbage, fennel and prebiotics such as fructooligosaccharides (FOS), oligofructose and inulin.[24][25] While many of these foods lack in butyrate production compared to resistant starch, they do have a number of benefits. They generally possess a low glycemic index which appeals well to diabetics. They also appeal to those on the ketogenic diet who benefit from beta-hydroxybutyric acid, which is a HDAC inhibitor which can cross the blood brain barrier and be used as fuel in the mitochondria of brain cells.[26] Other HDAC inhibitors in these butyrate producing foods are sulforaphane,[27] which has promise in inhibiting human breast cancer cells.[28] Sulforaphane has also been shown to promote hair growth in mice,[29] it contains compounds which prevent ulcers,[30] and helps with cognitive function in rats.[31] It is good to note that sulforaphane in broccoli is destroyed if prepared improperly.[32] Diallyl disulfide found in the fructans containing garlic has been shown to reduce chemical toxicity and carcinogenesis in rodents,[33] and shows synergestic benefits with butyrate when it comes to inhibiting the growth of human cancer tumor cells in the colon.[34]
Pharmacology
| Inhibited enzyme | IC50 (nM) | Entry note |
|---|---|---|
| HDAC1 | 16,000 | |
| HDAC2 | 12,000 | |
| HDAC3 | 9,000 | |
| HDAC4 | 2,000,000 | Lower bound |
| HDAC5 | 2,000,000 | Lower bound |
| HDAC6 | 2,000,000 | Lower bound |
| HDAC7 | 2,000,000 | Lower bound |
| HDAC8 | 15,000 | |
| HDAC9 | 2,000,000 | Lower bound |
| CA1 | 511,000 | |
| CA2 | 1,032,000 | |
| GPCR target | pEC50 | Entry note |
| FFAR2 | 2.9–4.6 | Full agonist |
| FFAR3 | 3.8–4.9 | Full agonist |
| NIACR1 | missing data | Full agonist |
Pharmacodynamics
Like other short chain fatty acids (SCFAs), butyrate is an agonist at the free fatty acid receptors FFAR2 and FFAR3, which function as nutrient sensors which help regulate energy balance;[37][38][39] unlike the other SCFAs,[39] butyrate is also an agonist of niacin receptor 1 (GPR109A).[37][38][39] Butyric acid is utilized by mitochondria, particularly in colonocytes and by the liver, to generate adenosine triphosphate (ATP) during fatty acid metabolism.[37] Butyric acid is also an HDAC inhibitor (specifically, HDAC1, HDAC2, HDAC3, and HDAC8),[35][36] a drug that inhibits the function of histone deacetylase enzymes, thereby favoring an acetylated state of histones in cells.[37] Histone acetylation loosens the structure of chromatin by reducing the electrostatic attraction between histones and DNA.[37] In general, it is thought that transcription factors will be unable to access regions where histones are tightly associated with DNA (i.e., non-acetylated, e.g., heterochromatin). Therefore, butyric acid is thought to enhance the transcriptional activity at promoters,[37] which are typically silenced or downregulated due to histone deacetylase activity.
Pharmacokinetics
Butyrate that is produced in the colon through microbial fermentation of dietary fiber is primarily absorbed and utilized by colonocytes and the liver[note 1] for the generation of ATP during energy metabolism;[37] however, some butyrate is absorbed in the distal colon, which is not connected to the portal vein, thereby allowing for the systemic distribution of butyrate to multiple organ systems through the circulatory system.[37] Butyrate that has reached systemic circulation can readily cross the blood-brain barrier via monocarboxylate transporters (i.e., certain members of the SLC16A group of transporters).[40][41] Other transporters that mediate the passage of butyrate across lipid membranes include SLC5A8 (SMCT1), SLC27A1 (FATP1), and SLC27A4 (FATP4).[35][41]
Metabolism
Butyric acid is metabolized by various human XM-ligases (ACSM1, ACSM2B, ASCM3, ACSM4, ACSM5, and ACSM6), also known as butyrate–CoA ligase.[10] The metabolite produced by this reaction is butyryl–CoA, and is produced as follows:[10]
- Adenosine triphosphate + Butyric acid + Coenzyme A → Adenosine monophosphate + Pyrophosphate + Butyryl-CoA
As a short chain fatty acid, butyrate is utilized by mitochondria as an energy (i.e., adenosine triphosphate or ATP) source through fatty acid metabolism.
In humans, the butyrate prodrug tributyrin is metabolized by triacylglycerol lipase into dibutyrin and butyrate through the reaction:[42]
- Tributyrin + H20 = Dibutyrin + Butyrate
Research
Peripheral therapeutic effects
Butyrate has numerous beneficial effects in humans on energy homeostasis and related diseases (diabetes and obesity), inflammation, and immune function (e.g., it has pronounced antimicrobial and anticarcinogenic effects).[38][43] These effects occur through its utilization by mitochondria to generate ATP during fatty acid metabolism or through one or more of its histone-modifying enzyme targets (i.e., the class I histone deacetylases) and G-protein coupled receptor targets (i.e., FFAR2, FFAR3, and NIACR1).[38]
Immunomodulation and inflammation
Butyrate's effects on the immune system are mediated through the inhibition of class I histone deacetylases and activation of its G-protein coupled receptor targets: NIACR1 (GPR109A), FFAR2 (GPR43), and FFAR3 (GPR41).[39][44] Among the short-chain fatty acids, butyrate is the most potent promoter of intestinal regulatory T cells in vitro and the only one among the group that is an NIACR1 ligand.[39] It has been shown to be a critical mediator of the colonic inflammatory response. It possesses both preventive and therapeutic potential to counteract inflammation-mediated ulcerative colitis and colorectal cancer.
Butyrate has established antimicrobial properties in humans that are mediated through the antimicrobial peptide LL-37, which it induces via HDAC inhibition on histone H3.[44][45][46] Butyrate increases gene expression of FOXP3 (the transcription regulator for Tregs) and promotes colonic regulatory T cells (Tregs) through the inhibition of class I histone deacetylases;[39][44] through these actions, it increases the expression of interleukin 10, an anti-inflammatory cytokine.[44][39] Butyrate also suppresses colonic inflammation by inhibiting the IFN-γ–STAT1 signaling pathways, which is mediated partially through histone deacetylase inhibition. While transient IFN-γ signaling is generally associated with normal host immune response, chronic IFN-γ signaling is often associated with chronic inflammation. It has been shown that butyrate inhibits activity of HDAC1 that is bound to the Fas gene promoter in T cells, resulting in hyperacetylation of the Fas promoter and up-regulation of Fas receptor on the T-cell surface.[47] It is thus suggested that butyrate enhances apoptosis of T cells in the colonic tissue and thereby eliminates the source of inflammation (IFN-γ production).[47]
Similar to other NIACR1 agonists, butyrate also produces marked anti-inflammatory effects in a variety of tissues, including the brain, gastrointestinal tract, skin, and vascular tissue.[48][49][50][51] Butyrate binding at FFAR3 induces neuropeptide Y release and promotes the functional homeostasis of colonic mucosa and the enteric immune system.[52]
Butyric acid is important as an energy (ATP) source for cells lining the mammalian colon (colonocytes). Without butyric acid for energy, colon cells undergo upregulated autophagy (i.e., self-digestion).[53]
Cancer
Butyrate produces different effects in healthy and cancerous cells; this is known as the "butyrate paradox". In particular, butyrate inhibits colonic tumor cells and promotes healthy colonic epithelial cells.[54] The signaling mechanism is not well understood.[55] A review suggested that the chemopreventive benefits of butyrate depend in part on the amount, time of exposure with respect to the tumorigenic process, and type of fat in the diet.[20] The production of volatile fatty acids such as butyrate from fermentable fibers may contribute to the role of dietary fiber in colon cancer.[20] Short-chain fatty acids, which include butyric acid, are produced by beneficial colonic bacteria (probiotics) that feed on, or ferment prebiotics, which are plant products that contain adequate amounts of dietary fiber. These short-chain fatty acids benefit the colonocytes by increasing energy production and cell proliferation, and may protect against colon cancer.[56]
Conversely, some researchers have sought to eliminate butyrate and consider it a potential cancer driver.[57] Studies in mice indicate it drives transformation of MSH2-deficient colon epithelial cells.[58] It is important to note that these are related to a gene deficiency. Niacin, beta-hydroxybutyrate, and curcumin may be effective adjunct treatments if genetic issues are present.[59]
Diabetes
A review on the relationship between the microbiome and diabetes asserted that butyrate can induce "profound immunometabolic effects" in animal models and humans with type 2 diabetes;[43] it also noted a relationship between the presence of obesity or diabetes and a state of marked dysbiosis in a host, which is not yet completely understood.[43] While acknowledging that there is strong evidence for the use of butyrate in such disorders, the review called for more research into the pathophysiology (i.e., biomolecular mechanisms) of these diseases, so as to improve therapeutic approaches to these diseases.[43]
Neuroepigenetic effects
Addiction
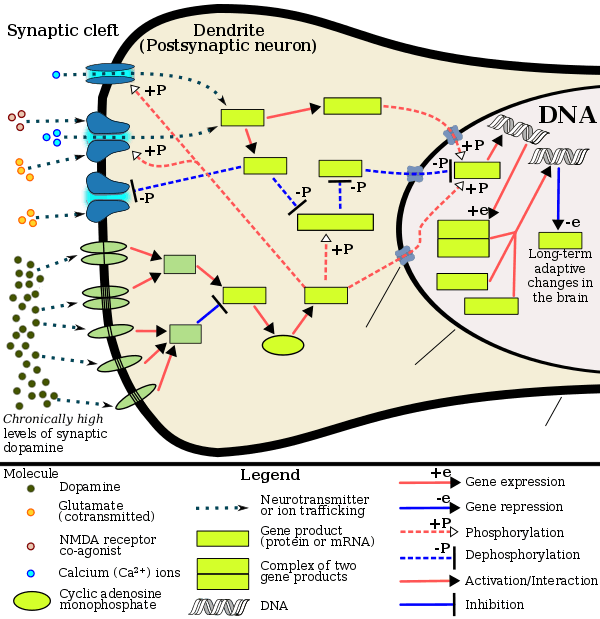
The observation of a large number of downregulated genes after methamphetamine withdrawal is consistent with previous results showing that methamphetamine can cause increased expression of histone deacetylases (HDACs) in the nucleus accumbens and the dorsal striatum. Butyric acid is a HDAC inhibitor.[67] HDACs are enzymes that can cause histone deacetylation and repression of gene expression. HDACs are important regulators of synaptic formation, synaptic plasticity, and long-term memory formation. Several HDACs also appear to play significant roles in various models of drug abuse and addiction.[68] The local knockout of HDAC1, as well as chronic and continuous infusion of MS-275, a pharmacological inhibitor highly selective in vitro for HDAC1, has been found with NAc suppressed cocaine-induced locomotor sensitization in mice.[69] HDAC3 inhibitor RGFP966 has been shown to facilitate the extinction of cocaine-seeking behavior and prevent reinstatement of cocaine-conditioned place preference in mice.[70] Histone deacetylase inhibitors have been shown to decrease cocaine, but not sucrose, self-administration in rats.[71] The beneficial bacteria that ferment probiotics and prebiotics to produce butyric acid have been shown to regulate behavior by means of the vagus nerve.[72]
Cognitive deficits and memory
Studies in rodents have found that the environment exerts an influence on epigenetic changes related to cognition, in terms of learning and memory;[73] environmental enrichment is correlated with increased histone acetylation, and verification by administering histone deacetylase inhibitors induced the sprouting of dendrites, an increased number of synapses, and reinstated learning behaviour and access to long-term memories.[74][75] Research has also linked learning and long-term memory formation to reversible epigenetic changes in the hippocampus and cortex in animals with normal-functioning, undamaged brains.[75][76] In human studies, post-mortem brains from Alzheimer's patients show increased histone de-acetylase levels.[77][78]
See also
- Category:Butyrates
- Histone
- Histone-modifying enzyme
- Hydroxybutyric acids
- β-Hydroxy β-methylbutyric acid
- β-Methylbutyric acid
- Synbiotics
Notes
- ↑ Most of the butyrate that is absorbed into blood plasma from the colon enters the circulatory system via the portal vein;[37] most of the butyrate that enters the circulatory system by this route is taken up by the liver.[37]
References
![]() This article incorporates text from a publication now in the public domain: Chisholm, Hugh, ed. (1911). "Butyric Acid". Encyclopædia Britannica (11th ed.). Cambridge University Press.
This article incorporates text from a publication now in the public domain: Chisholm, Hugh, ed. (1911). "Butyric Acid". Encyclopædia Britannica (11th ed.). Cambridge University Press.
- 1 2 Nomenclature of Organic Chemistry : IUPAC Recommendations and Preferred Names 2013 (Blue Book). Cambridge: The Royal Society of Chemistry. 2014. p. 746. doi:10.1039/9781849733069-FP001. ISBN 978-0-85404-182-4.
- 1 2 3 4 Strieter FJ, Templeton DH (1962). "Crystal structure of butyric acid". Acta Crystallographica. 15 (12): 1240–1244. doi:10.1107/S0365110X6200328X.
- 1 2 3 4 Lide, David R., ed. (2009). CRC Handbook of Chemistry and Physics (90th ed.). Boca Raton, Florida: CRC Press. ISBN 978-1-4200-9084-0.
- 1 2 3 4 5 Butanoic acid in Linstrom, P.J.; Mallard, W.G. (eds.) NIST Chemistry WebBook, NIST Standard Reference Database Number 69. National Institute of Standards and Technology, Gaithersburg MD. http://webbook.nist.gov (retrieved 13 June 2014)
- 1 2 3 4 5 6 CID 264 from PubChem
- 1 2 3 4 "Butanoic Acid". ALS Environmental. Retrieved 13 June 2014.
- 1 2 3 "butanoic acid". Chemister.ru. 2007-03-19. Retrieved 2016-05-09.
- 1 2 3 4 5 Sigma-Aldrich Co., Butyric acid. Retrieved on 13 June 2014.
- ↑ Moskin, Julia (13 February 2008). "Dark may be king, but milk chocolate makes a move". The New York Times. Retrieved 1 January 2016.
- 1 2 3 "Butyric acid". Human Metabolome Database. University of Alberta. Retrieved 15 August 2015.
- ↑ Chevreul (1815) "Lettre de M. Chevreul à MM. les rédacteurs des Annales de chimie" (Letter from Mr. Chevreul to the editors of the Annals of Chemistry), Annales de chimie, 94 : 73–79; in a footnote spanning pages 75–76, he mentions that he had found a substance that is responsible for the smell of butter.
- ↑ Chevreul (1817) "Extrait d'une lettre de M. Chevreul à MM. les Rédacteurs du Journal de Pharmacie" (Extract of a letter from Mr. Chevreul to the editors of the Journal of Pharmacy), Journal de Pharmacie et des sciences accessoires, 3 : 79–81. On p. 81, he named butyric acid: "Ce principe, que j'ai appelé depuis acid butérique, … " (This principle [i.e., constituent], which I have since named "butyric acid", … )
- ↑ E. Chevreul, Recherches chimiques sur les corps gras d'origine animale [Chemical researches on fatty substances of animal origin] (Paris, France: F.G. Levrault, 1823), pages 115–133.
- 1 2 ICSC 1334 – BUTYRIC ACID. Inchem.org (23 November 1998). Retrieved on 2014-03-31.
- ↑ Supplementation of Coated Butyric Acid in the Feed Reduces Colonization and Shedding of Salmonella in Poultry. Ps.fass.org. Retrieved on 31 March 2014.
- ↑ Freezer Baits, nutrabaits.net
- ↑ Kasumyan A, Døving K (2003). "Taste preferences in fishes". Fish and Fisheries. 4 (4): 289–347. doi:10.1046/j.1467-2979.2003.00121.x.
- ↑ Japanese Whalers Injured by Acid-Firing Activists, newser.com, 10 February 2010
- ↑ National Abortion Federation, HISTORY OF VIOLENCE Butyric Acid Attacks. Prochoice.org. Retrieved on 31 March 2014.
- 1 2 3 Lupton JR (February 2004). "Microbial degradation products influence colon cancer risk: the butyrate controversy". The Journal of Nutrition. 134 (2): 479–82. PMID 14747692.
- ↑ Cummings JH, Macfarlane GT, Englyst HN (February 2001). "Prebiotic digestion and fermentation". The American Journal of Clinical Nutrition. 73 (2 Suppl): 415S–420S. PMID 11157351.
- ↑ Grummer RR (September 1991). "Effect of feed on the composition of milk fat" (PDF). Journal of Dairy Science. 74 (9): 3244–57. doi:10.3168/jds.S0022-0302(91)78510-X. PMID 1779073.
- ↑ webmed. "Frequently asked questions in the area of diet and IBS". www.med.monash.edu. Retrieved 24 March 2016.
- ↑ Gibson, Peter R.; Shepherd, Susan J. (1 February 2010). "Evidence-based dietary management of functional gastrointestinal symptoms: The FODMAP approach". Journal of Gastroenterology and Hepatology. 25 (2): 252–258. doi:10.1111/j.1440-1746.2009.06149.x. ISSN 1440-1746. PMID 20136989.
- ↑ Gibson, Peter R.; Varney, Jane; Malakar, Sreepurna; Muir, Jane G. (1 May 2015). "Food components and irritable bowel syndrome". Gastroenterology. 148 (6): 1158–1174.e4. doi:10.1053/j.gastro.2015.02.005. ISSN 1528-0012. PMID 25680668.
- ↑ Owen, O. E.; Morgan, A. P.; Kemp, H. G.; Sullivan, J. M.; Herrera, M. G.; Cahill, G. F. (1 October 1967). "Brain metabolism during fasting". The Journal of Clinical Investigation. 46 (10): 1589–1595. doi:10.1172/JCI105650. ISSN 0021-9738. PMC 292907
 . PMID 6061736.
. PMID 6061736. - ↑ Ho, Emily; Clarke, John D.; Dashwood, Roderick H. (1 December 2009). "Dietary Sulforaphane, a Histone Deacetylase Inhibitor for Cancer Prevention". The Journal of Nutrition. 139 (12): 2393–2396. doi:10.3945/jn.109.113332. ISSN 0022-3166. PMC 2777483. PMID 19812222.
- ↑ Pledgie-Tracy, Allison; Sobolewski, Michele D.; Davidson, Nancy E. (1 March 2007). "Sulforaphane induces cell type–specific apoptosis in human breast cancer cell lines". Molecular Cancer Therapeutics. 6 (3): 1013–1021. doi:10.1158/1535-7163.MCT-06-0494. ISSN 1535-7163. PMID 17339367.
- ↑ Sasaki, Mari; Shinozaki, Shohei; Shimokado, Kentaro (25 March 2016). "Sulforaphane promotes murine hair growth by accelerating the degradation of dihydrotestosterone". Biochemical and Biophysical Research Communications. 472 (1): 250–254. doi:10.1016/j.bbrc.2016.02.099. ISSN 1090-2104. PMID 26923074.
- ↑ Moon, Joon-Kwan; Kim, Jun-Ran; Ahn, Young-Joon; Shibamoto, Takayuki (9 June 2010). "Analysis and anti-Helicobacter activity of sulforaphane and related compounds present in broccoli ( Brassica oleracea L.) sprouts". Journal of Agricultural and Food Chemistry. 58 (11): 6672–6677. doi:10.1021/jf1003573. ISSN 1520-5118. PMID 20459098.
- ↑ Dash, Pramod K.; Zhao, Jing; Orsi, Sara A.; Zhang, Min; Moore, Anthony N. (28 August 2009). "Sulforaphane improves cognitive function administered following traumatic brain injury". Neuroscience Letters. 460 (2): 103–107. doi:10.1016/j.neulet.2009.04.028. ISSN 1872-7972. PMC 2700200. PMID 19515491.
- ↑ Ghawi, Sameer Khalil; Methven, Lisa; Niranjan, Keshavan. "The potential to intensify sulforaphane formation in cooked broccoli (Brassica oleracea var. italica) using mustard seeds (Sinapis alba)". Food Chemistry. 138 (2–3): 1734–1741. doi:10.1016/j.foodchem.2012.10.119.
- ↑ Yang, C. S.; Chhabra, S. K.; Hong, J. Y.; Smith, T. J. (1 March 2001). "Mechanisms of inhibition of chemical toxicity and carcinogenesis by diallyl sulfide (DAS) and related compounds from garlic". The Journal of Nutrition. 131 (3s): 1041S–5S. ISSN 0022-3166. PMID 11238812.
- ↑ Sundaram, Sujatha G.; Milner, John A. (1 April 1996). "Diallyl disulfide induces apoptosis of human colon tumor cells". Carcinogenesis. 17 (4): 669–673. doi:10.1093/carcin/17.4.669. ISSN 0143-3334. PMID 8625476.
- 1 2 3 "Butyric acid". IUPHAR. IUPHAR/BPS Guide to PHARMACOLOGY. Retrieved 23 May 2015.
- 1 2 "butanoic acid, 4 and Sodium; butyrate". BindingDB. The Binding Database. Retrieved 23 May 2015.
- 1 2 3 4 5 6 7 8 9 10 11 Bourassa MW, Alim I, Bultman SJ, Ratan RR (June 2016). "Butyrate, neuroepigenetics and the gut microbiome: Can a high fiber diet improve brain health?". Neurosci. Lett. 625: 56–63. doi:10.1016/j.neulet.2016.02.009. PMID 26868600.
Butyrate is an attractive therapeutic molecule because of its wide array of biological functions, such as its ability to serve as a histone deacetylase (HDAC) inhibitor, an energy metabolite to produce ATP and a G protein-coupled receptor (GPCR) activator. ... Histone acetylation is a post-translational modification by an epigenetic protein, which are proteins that bind to chromatin and influence chromatin structure to change the propensity that a gene is transcribed or repressed. Acetylated histones cause the chromatin structure to loosen by weakening electrostatic attraction between the histone proteins and the DNA backbone. This process enables transcription factors and the basal transcriptional machinery to bind and increases transcription. ... However, many studies have shown that at least some of these beneficial effects can be attributed NaB’s ability to increase acetylation around the promoters of neurotrophic factors, such as BDNF, GDNF and NGF and thus increasing their transcription [41], [42], [43], [44], [45], [46], [47] and [48]. ... Butyrate also signals through GPR109a ... Much of the butyrate produced in the colon is used as an energy source by the colonocytes, but some butyrate can also exit the colon through the portal vein, where the liver absorbs another large portion [74] and [75]. However, the distal colon is not connected to the portal vein, allowing for some systemic butyrate to be circulated. Indeed, there are many reports of high fiber diets increasing blood levels of circulating butyrate [75], [76] and [77]. These later reports raise the possibility that increases in circulating butyrate could affect CNS function directly.
- 1 2 3 4 Kasubuchi M, Hasegawa S, Hiramatsu T, Ichimura A, Kimura I (2015). "Dietary gut microbial metabolites, short-chain fatty acids, and host metabolic regulation". Nutrients. 7 (4): 2839–49. doi:10.3390/nu7042839. PMC 4425176. PMID 25875123.
Short-chain fatty acids (SCFAs) such as acetate, butyrate, and propionate, which are produced by gut microbial fermentation of dietary fiber, are recognized as essential host energy sources and act as signal transduction molecules via G-protein coupled receptors (FFAR2, FFAR3, OLFR78, GPR109A) and as epigenetic regulators of gene expression by the inhibition of histone deacetylase (HDAC). Recent evidence suggests that dietary fiber and the gut microbial-derived SCFAs exert multiple beneficial effects on the host energy metabolism not only by improving the intestinal environment, but also by directly affecting various host peripheral tissues.
- 1 2 3 4 5 6 7 Hoeppli RE, Wu D, Cook L, Levings MK (February 2015). "The environment of regulatory T cell biology: cytokines, metabolites, and the microbiome". Front Immunol. 6: 61. doi:10.3389/fimmu.2015.00061. PMC 4332351. PMID 25741338.
Specific species that have been recognized by their high levels of butyrate production include Faecalibacterium prausnitzii and the cluster IV and XIVa of genus Clostridium ... Administration of acetate, propionate, and butyrate in drinking water mimics the effect of Clostridium colonization in germ-free mice, resulting in an elevated Treg frequency in the colonic lamina propria and increased IL-10 production by these Tregs (180, 182). Of the three main SCFAs, butyrate has been found to be the most potent inducer of colonic Tregs. Mice fed a diet enriched in butyrylated starches have more colonic Tregs than those fed a diet containing propinylated or acetylated starches (181). Arpaia et al. tested an array of SCFAs purified from commensal bacteria and confirmed butyrate was the strongest SCFA-inducer of Tregs in vitro (180). Mechanistically, it has been proposed that butyrate, and possibly propionate, promote Tregs through inhibiting histone deacetylase (HDAC), causing increased acetylation of histone H3 in the Foxp3 CNS1 region, and thereby enhancing FOXP3 expression (180, 181). Short-chain fatty acids partially mediate their effects through G-protein coupled receptors (GPR), including GPR41, GPR43, and GPR109A. GPR41 and GPR43 are stimulated by all three major SCFAs (191), whereas GPR109A only interacts with butyrate (192).
Figure 1: Microbial-derived molecules promote colonic Treg differentiation. - ↑ Tsuji A (2005). "Small molecular drug transfer across the blood-brain barrier via carrier-mediated transport systems". NeuroRx. 2 (1): 54–62. doi:10.1602/neurorx.2.1.54. PMC 539320. PMID 15717057.
Other in vivo studies in our laboratories indicated that several compounds including acetate, propionate, butyrate, benzoic acid, salicylic acid, nicotinic acid, and some β-lactam antibiotics may be transported by the MCT at the BBB.21 ... Uptake of valproic acid was reduced in the presence of medium-chain fatty acids such as hexanoate, octanoate, and decanoate, but not propionate or butyrate, indicating that valproic acid is taken up into the brain via a transport system for medium-chain fatty acids, not short-chain fatty acids.
- 1 2 Vijay N, Morris ME (2014). "Role of monocarboxylate transporters in drug delivery to the brain". Curr. Pharm. Des. 20 (10): 1487–98. doi:10.2174/13816128113199990462. PMC 4084603. PMID 23789956.
Monocarboxylate transporters (MCTs) are known to mediate the transport of short chain monocarboxylates such as lactate, pyruvate and butyrate. ... MCT1 and MCT4 have also been associated with the transport of short chain fatty acids such as acetate and formate which are then metabolized in the astrocytes [78]. ... SLC5A8 is expressed in normal colon tissue, and it functions as a tumor suppressor in human colon with silencing of this gene occurring in colon carcinoma. This transporter is involved in the concentrative uptake of butyrate and pyruvate produced as a product of fermentation by colonic bacteria.
- ↑ "triacylglycerol lipase – Homo sapiens". BRENDA. Technische Universität Braunschweig. Retrieved 25 May 2015.
- 1 2 3 4 Tilg H, Moschen AR (September 2014). "Microbiota and diabetes: an evolving relationship". Gut. 63 (9): 1513–1521. doi:10.1136/gutjnl-2014-306928. PMID 24833634.
Recent studies have suggested that gut bacteria play a fundamental role in diseases such as obesity, diabetes and cardiovascular disease. Data are accumulating in animal models and humans suggesting that obesity and type 2 diabetes (T2D) are associated with a profound dysbiosis. First human metagenome-wide association studies demonstrated highly significant correlations of specific intestinal bacteria, certain bacterial genes and respective metabolic pathways with T2D. Importantly, especially butyrate-producing bacteria such as Roseburia intestinalis and Faecalibacterium prausnitzii concentrations were lower in T2D subjects. This supports the increasing evidence, that butyrate and other short-chain fatty acids are able to exert profound immunometabolic effects.
- 1 2 3 4 Wang G (2014). "Human antimicrobial peptides and proteins". Pharmaceuticals (Basel). 7 (5): 545–94. doi:10.3390/ph7050545. PMC 4035769. PMID 24828484.
The establishment of a link between light therapy, vitamin D and human cathelicidin LL-37 expression provides a completely different way for infection treatment. Instead of treating patients with traditional antibiotics, doctors may be able to use light or vitamin D [291,292]. Indeed using narrow-band UV B light, the level of vitamin D was increased in psoriasis patients (psoriasis is a common autoimmune disease on skin) [293]. In addition, other small molecules such as butyrate can induce LL-37 expression [294]. Components from Traditional Chinese Medicine may regulate the AMP expression as well [295]. These factors may induce the expression of a single peptide or multiple AMPs [296]. It is also possible that certain factors can work together to induce AMP expression. While cyclic AMP and butyrate synergistically stimulate the expression of chicken β-defensin 9 [297], 4-phenylbutyrate (PBA) and 1,25-dihydroxyvitamin D3 (or lactose) can induce AMP gene expression synergistically [294,298]. It appears that stimulation of LL-37 expression by histone deacetylase (HDAC) inhibitors is cell dependent. Trichostatin and sodium butyrate increased the peptide expression in human NCI-H292 airway epithelial cells but not in the primary cultures of normal nasal epithelial cells [299]. However, the induction of the human LL-37 expression may not be a general approach for bacterial clearance. During Salmonella enterica infection of human monocyte-derived macrophages, LL-37 is neither induced nor required for bacterial clearance [300].
Table 3: Select human antimicrobial peptides and their proposed targets
Table 4: Some known factors that induce antimicrobial peptide expression - ↑ Yonezawa H, Osaki T, Hanawa T, Kurata S, Zaman C, Woo TD, Takahashi M, Matsubara S, Kawakami H, Ochiai K, Kamiya S (2012). "Destructive effects of butyrate on the cell envelope of Helicobacter pylori". J. Med. Microbiol. 61 (Pt 4): 582–9. doi:10.1099/jmm.0.039040-0. PMID 22194341.
- ↑ McGee DJ, George AE, Trainor EA, Horton KE, Hildebrandt E, Testerman TL (2011). "Cholesterol enhances Helicobacter pylori resistance to antibiotics and LL-37". Antimicrob. Agents Chemother. 55 (6): 2897–904. doi:10.1128/AAC.00016-11. PMC 3101455. PMID 21464244.
- 1 2 Zimmerman MA, Singh N, Martin PM, Thangaraju M, Ganapathy V, Waller JL, Shi H, Robertson KD, Munn DH, Liu K (2012). "Butyrate suppresses colonic inflammation through HDAC1-dependent Fas upregulation and Fas-mediated apoptosis of T cells". Am. J. Physiol. Gastrointest. Liver Physiol. 302 (12): G1405–15. doi:10.1152/ajpgi.00543.2011. PMC 3378095. PMID 22517765.
- ↑ Offermanns S, Schwaninger M (2015). "Nutritional or pharmacological activation of HCA(2) ameliorates neuroinflammation". Trends Mol Med. 21 (4): 245–255. doi:10.1016/j.molmed.2015.02.002. PMID 25766751.
Neuroinflammatory cells express HCA2, a receptor for the endogenous neuroprotective ketone body β-hydroxybutyrate (BHB) as well as for the drugs dimethyl fumarate (DMF) and nicotinic acid, which have established efficacy in the treatment of MS and experimental stroke, respectively. This review summarizes the evidence that HCA2 is involved in the therapeutic effects of DMF, nicotinic acid, and ketone bodies in reducing neuroinflammation.
- ↑ Chai JT, Digby JE, Choudhury RP (May 2013). "GPR109A and vascular inflammation". Curr Atheroscler Rep. 15 (5): 325. doi:10.1007/s11883-013-0325-9. PMC 3631117. PMID 23526298.
As GPR109A's primary pharmacological ligand in clinical use, niacin has been used for over 50 years in the treatment of cardiovascular disease, mainly due to its favourable effects on plasma lipoproteins. However, it has become apparent that niacin also possesses lipoprotein-independent effects that influence inflammatory pathways mediated through GPR109A.
- ↑ Graff EC, Fang H, Wanders D, Judd RL (February 2016). "Anti-inflammatory effects of the hydroxycarboxylic acid receptor 2". Metab. Clin. Exp. 65 (2): 102–113. doi:10.1016/j.metabol.2015.10.001. PMID 26773933.
HCA2 is highly expressed on immune cells, including macrophages, monocytes, neutrophils and dermal dendritic cells, among other cell types. ... Recent studies demonstrate that HCA2 mediates profound anti-inflammatory effects in a variety of tissues, indicating that HCA2 may be an important therapeutic target for treating inflammatory disease processes.
- ↑ Wakade C, Chong R (December 2014). "A novel treatment target for Parkinson's disease". J. Neurol. Sci. 347 (1-2): 34–38. doi:10.1016/j.jns.2014.10.024. PMID 25455298.
GPR109A and its agonists are known to exert anti-inflammatory actions in the skin, gut and retina.
- ↑ Farzi A, Reichmann F, Holzer P (2015). "The homeostatic role of neuropeptide Y in immune function and its impact on mood and behaviour". Acta Physiol (Oxf). 213 (3): 603–27. doi:10.1111/apha.12445. PMC 4353849. PMID 25545642.
In the context of this review it is particularly worth noting that short chain fatty acids such as butyrate, which the colonic microbiota generates by fermentation of otherwise indigestible dietary fibre (Cherbut et al. 1998), stimulate L cells to release PYY via the G-protein coupled receptor Gpr41 (Samuel et al. 2008). In this way, short chain fatty acids can indirectly attenuate gastrointestinal motility as well as electrolyte and water secretion (Cox 2007b). More importantly, short chain fatty acids exert homeostatic actions on the function of the colonic mucosa and immune system (Hamer et al. 2008, Tazoe et al. 2008, Guilloteau et al. 2010, Macia et al. 2012a, Smith et al. 2013). Whether PYY plays a role in these effects of short chain fatty acids awaits to be investigated, but may be envisaged from the finding that PYY promotes mucosal cell differentiation (Hallden & Aponte 1997).
- ↑ Donohoe, Dallas R.; Garge, Nikhil; Zhang, Xinxin; Sun, Wei; O’Connell, Thomas M.; Bunger, Maureen K.; Bultman, Scott J. (4 May 2011). "The Microbiome and Butyrate Regulate Energy Metabolism and Autophagy in the Mammalian Colon". Cell Metabolism. 13 (5): 517–526. doi:10.1016/j.cmet.2011.02.018. ISSN 1550-4131. PMC 3099420. PMID 21531334.
- ↑ Vanhoutvin SA, Troost FJ, Hamer HM, Lindsey PJ, Koek GH, Jonkers DM, Kodde A, Venema K, Brummer RJ (2009). Bereswill S, ed. "Butyrate-induced transcriptional changes in human colonic mucosa". PLOS ONE. 4 (8): e6759. doi:10.1371/journal.pone.0006759. PMC 2727000. PMID 19707587.
- ↑ Klampfer L, Huang J, Sasazuki T, Shirasawa S, Augenlicht L (August 2004). "Oncogenic Ras promotes butyrate-induced apoptosis through inhibition of gelsolin expression" (PDF). The Journal of Biological Chemistry. 279 (35): 36680–8. doi:10.1074/jbc.M405197200. PMID 15213223.
- ↑ Lupton, Joanne R. (2004). Microbial Degradation Products Influence Colon Cancer Risk: the Butyrate Controversy. vol. 134 no. 2: J. Nutr. pp. 479–482.
- ↑ "Low-carb diet cuts risk of colon cancer, study finds | University of Toronto Media Room". media.utoronto.ca. Retrieved 2016-05-04.
- ↑ Belcheva, Antoaneta; Irrazabal, Thergiory; Robertson, Susan J.; Streutker, Catherine; Maughan, Heather; Rubino, Stephen; Moriyama, Eduardo H.; Copeland, Julia K.; Kumar, Sachin (2014-07-17). "Gut microbial metabolism drives transformation of MSH2-deficient colon epithelial cells". Cell. 158 (2): 288–299. doi:10.1016/j.cell.2014.04.051. ISSN 1097-4172. PMID 25036629.
- ↑ "Colon Cancer Shuts Down Receptor That Could Shut It Down". ScienceDaily. Retrieved 2016-05-04.
- 1 2 3 Renthal W, Nestler EJ (September 2009). "Chromatin regulation in drug addiction and depression". Dialogues Clin. Neurosci. 11 (3): 257–268. PMC 2834246. PMID 19877494. Retrieved 21 July 2014.
[Psychostimulants] increase cAMP levels in striatum, which activates protein kinase A (PKA) and leads to phosphorylation of its targets. This includes the cAMP response element binding protein (CREB), the phosphorylation of which induces its association with the histone acetyltransferase, CREB binding protein (CBP) to acetylate histones and facilitate gene activation. This is known to occur on many genes including fosB and c-fos in response to psychostimulant exposure. ΔFosB is also upregulated by chronic psychostimulant treatments, and is known to activate certain genes (eg, cdk5) and repress others (eg, c-fos) where it recruits HDAC1 as a corepressor. ... Chronic exposure to psychostimulants increases glutamatergic [signaling] from the prefrontal cortex to the NAc. Glutamatergic signaling elevates Ca2+ levels in NAc postsynaptic elements where it activates CaMK (calcium/calmodulin protein kinases) signaling, which, in addition to phosphorylating CREB, also phosphorylates HDAC5.
- ↑ Broussard JI (January 2012). "Co-transmission of dopamine and glutamate". J. Gen. Physiol. 139 (1): 93–96. doi:10.1085/jgp.201110659. PMC 3250102. PMID 22200950.
- ↑ Kanehisa Laboratories (10 October 2014). "Amphetamine – Homo sapiens (human)". KEGG Pathway. Retrieved 31 October 2014.
Most addictive drugs increase extracellular concentrations of dopamine (DA) in nucleus accumbens (NAc) and medial prefrontal cortex (mPFC), projection areas of mesocorticolimbic DA neurons and key components of the "brain reward circuit". Amphetamine achieves this elevation in extracellular levels of DA by promoting efflux from synaptic terminals. ... Chronic exposure to amphetamine induces a unique transcription factor delta FosB, which plays an essential role in long-term adaptive changes in the brain.
- ↑ Cadet JL, Brannock C, Jayanthi S, Krasnova IN (2015). "Transcriptional and epigenetic substrates of methamphetamine addiction and withdrawal: evidence from a long-access self-administration model in the rat". Mol. Neurobiol. 51 (2): 696–717. doi:10.1007/s12035-014-8776-8. PMC 4359351. PMID 24939695.
Figure 1
- 1 2 3 Robison AJ, Nestler EJ (November 2011). "Transcriptional and epigenetic mechanisms of addiction". Nat. Rev. Neurosci. 12 (11): 623–637. doi:10.1038/nrn3111. PMC 3272277. PMID 21989194.
ΔFosB serves as one of the master control proteins governing this structural plasticity. ... ΔFosB also represses G9a expression, leading to reduced repressive histone methylation at the cdk5 gene. The net result is gene activation and increased CDK5 expression. ... In contrast, ΔFosB binds to the c-fos gene and recruits several co-repressors, including HDAC1 (histone deacetylase 1) and SIRT 1 (sirtuin 1). ... The net result is c-fos gene repression.
Figure 4: Epigenetic basis of drug regulation of gene expression - 1 2 3 Nestler EJ (December 2012). "Transcriptional mechanisms of drug addiction". Clin. Psychopharmacol. Neurosci. 10 (3): 136–143. doi:10.9758/cpn.2012.10.3.136. PMC 3569166. PMID 23430970.
The 35-37 kD ΔFosB isoforms accumulate with chronic drug exposure due to their extraordinarily long half-lives. ... As a result of its stability, the ΔFosB protein persists in neurons for at least several weeks after cessation of drug exposure. ... ΔFosB overexpression in nucleus accumbens induces NFκB ... In contrast, the ability of ΔFosB to repress the c-Fos gene occurs in concert with the recruitment of a histone deacetylase and presumably several other repressive proteins such as a repressive histone methyltransferase
- ↑ Nestler EJ (October 2008). "Review. Transcriptional mechanisms of addiction: role of DeltaFosB". Philos. Trans. R. Soc. Lond., B, Biol. Sci. 363 (1507): 3245–3255. doi:10.1098/rstb.2008.0067. PMC 2607320. PMID 18640924.
Recent evidence has shown that ΔFosB also represses the c-fos gene that helps create the molecular switch—from the induction of several short-lived Fos family proteins after acute drug exposure to the predominant accumulation of ΔFosB after chronic drug exposure
- ↑ Davie, James R. (2003-07-01). "Inhibition of Histone Deacetylase Activity by Butyrate". The Journal of Nutrition. 133 (7): 2485S–2493S. ISSN 0022-3166. PMID 12840228.
- ↑ Cadet, Jean Lud; Brannock, Christie; Jayanthi, Subramaniam; Krasnova, Irina N. (1 January 2015). "Transcriptional and Epigenetic Substrates of Methamphetamine Addiction and Withdrawal: Evidence from a Long-Access Self-Administration Model in the Rat". Molecular Neurobiology. 51 (2): 696–717. doi:10.1007/s12035-014-8776-8. ISSN 0893-7648. PMC 4359351. PMID 24939695.
- ↑ Kennedy, Pamela J.; Feng, Jian; Robison, A.J.; Maze, Ian; Badimon, Ana; Mouzon, Ezekiell; Chaudhury, Dipesh; Damez-Werno, Diane M.; Haggarty, Stephen J. (1 April 2013). "Class I HDAC Inhibition Blocks Cocaine-Induced Plasticity Through Targeted Changes in Histone Methylation". Nature Neuroscience. 16 (4): 434–440. doi:10.1038/nn.3354. ISSN 1097-6256. PMC 3609040. PMID 23475113.
- ↑ Malvaez, Melissa; McQuown, Susan C.; Rogge, George A.; Astarabadi, Mariam; Jacques, Vincent; Carreiro, Samantha; Rusche, James R.; Wood, Marcelo A. (12 February 2013). "HDAC3-selective inhibitor enhances extinction of cocaine-seeking behavior in a persistent manner". Proceedings of the National Academy of Sciences of the United States of America. 110 (7): 2647–2652. doi:10.1073/pnas.1213364110. ISSN 0027-8424. PMC 3574934. PMID 23297220.
- ↑ Romieu, Pascal; Host, Lionel; Gobaille, Serge; Sandner, Guy; Aunis, Dominique; Zwiller, Jean (17 September 2008). "Histone deacetylase inhibitors decrease cocaine but not sucrose self-administration in rats". The Journal of Neuroscience: The Official Journal of the Society for Neuroscience. 28 (38): 9342–9348. doi:10.1523/JNEUROSCI.0379-08.2008. ISSN 1529-2401. PMID 18799668.
- ↑ Bravo, Javier A.; Forsythe, Paul; Chew, Marianne V.; Escaravage, Emily; Savignac, Hélène M.; Dinan, Timothy G.; Bienenstock, John; Cryan, John F. (2011-09-20). "Ingestion of Lactobacillus strain regulates emotional behavior and central GABA receptor expression in a mouse via the vagus nerve". Proceedings of the National Academy of Sciences. 108 (38): 16050–16055. doi:10.1073/pnas.1102999108. ISSN 0027-8424. PMC 3179073. PMID 21876150.
- ↑ The Developing Genome: An Introduction to Behavioral Epigenetics (1 ed.). Oxford University Press. 2 March 2015. ISBN 9780199922345.
- ↑ Fischer, Andre; Sananbenesi, Farahnaz; Wang, Xinyu; Dobbin, Matthew; Tsai, Li-Huei (10 May 2007). "Recovery of learning and memory is associated with chromatin remodelling". Nature. 447 (7141): 178–182. doi:10.1038/nature05772. ISSN 1476-4687. PMID 17468743.
- 1 2 Miller, Greg (2 July 2010). "Epigenetics. The seductive allure of behavioral epigenetics". Science. 329 (5987): 24–27. doi:10.1126/science.329.5987.24. ISSN 1095-9203. PMID 20595592.
- ↑ Gupta, Swati; Kim, Se Y.; Artis, Sonja; Molfese, David L.; Schumacher, Armin; Sweatt, J. David; Paylor, Richard E.; Lubin, Farah D. (10 March 2010). "Histone methylation regulates memory formation". The Journal of Neuroscience: The Official Journal of the Society for Neuroscience. 30 (10): 3589–3599. doi:10.1523/JNEUROSCI.3732-09.2010. ISSN 1529-2401. PMC 2859898. PMID 20219993.
- ↑ Peleg, Shahaf; Sananbenesi, Farahnaz; Zovoilis, Athanasios; Burkhardt, Susanne; Bahari-Javan, Sanaz; Agis-Balboa, Roberto Carlos; Cota, Perla; Wittnam, Jessica Lee; Gogol-Doering, Andreas (7 May 2010). "Altered histone acetylation is associated with age-dependent memory impairment in mice". Science. 328 (5979): 753–756. doi:10.1126/science.1186088. ISSN 1095-9203. PMID 20448184.
- ↑ Gräff, Johannes; Rei, Damien; Guan, Ji-Song; Wang, Wen-Yuan; Seo, Jinsoo; Hennig, Krista M.; Nieland, Thomas J. F.; Fass, Daniel M.; Kao, Patricia F. (8 March 2012). "An epigenetic blockade of cognitive functions in the neurodegenerating brain". Nature. 483 (7388): 222–226. doi:10.1038/nature10849. ISSN 1476-4687. PMC 3498952. PMID 22388814.
External links
| Wikimedia Commons has media related to Butyric acid. |
- International Chemical Safety Card 1334
- 2004 review of the scientific evidence on butanoate/butyrate vs. colon cancer